
Samenvatting
De relatie tussen trauma, dissociatie en geheugen is voorwerp van intensieve studie geworden. Neurobiologisch onderzoek kan helpen deze relatie te verduidelijken. Traumatische stress resulteert in veranderingen in de hersenregionen die betrokken zijn bij de geheugenfuncties en de stressrespons; veranderingen hierin kunnen bij symptomen van dissociatie en de posttraumatische stress stoornis een rol spelen. Neuro-endocriene en neuro-anatomische limbische systemen worden beschreven als centrale elementen bij de integratie van declaratieve en impliciete geheugenfuncties. Bij de effecten van stress op geheugenfuncties zijn neurale mechanismen als angstconditionering, stress-sensitisatie en uitdoving betrokken. Het concept ‘neurale informatieverwerkende systemen’ ondersteunt de gedachte dat dissociatieve fenomenen in hoge mate ‘toestandsafhankelijk’ zijn. Het feit dat er bij traumatische herinneringen plastische veranderingen met neuro-endocriene parameters vastgesteld zijn maakt eenzelfde betekenis bij fictieve of geïmplanteerde herinneringen onwaarschijnlijk. Mogelijke aangrijpingspunten voor de toepassing van anti-dissociatieve farmaca worden beschreven.
Inleiding
De belangstelling voor de relaties tussen trauma, dissociatie en geheugen bestaat niet alleen bij de psychiatrische, psychologische en psychotherapeutische disciplines, maar ook bij neurobiologen (LeDoux, 1996; Schacter, 1996, Ederlyi, 1996; Conway, 1997; Bremner et al., 1995c; Bremner et al., 1997a, 1997b; Cahill, 1997). Twee verschijnselen krijgen bijzondere aandacht: enerzijds het onvermogen te vergeten – zoals zich dit bij bijvoorbeeld PTSS in de vorm van herbelevingen manifesteert; anderzijds het onvermogen om zich bepaalde aspecten van de traumatische ervaring of de ervaring als geheel te herinneren. Verschillende hypothesen zijn geopperd om deze ogenschijnlijk tegenstrijdige bevindingen te verklaren (zie bijvoorbeeld; Friedman et al, 1995; Crombag & Merckelbach, 1996; Draijer, 1997, Spinhoven, Nijenhuis & Van Dyck, 1998). Dissociatieve amnesie, ofwel het onvermogen zich belangrijke gebeurtenissen uit het eigen leven te herinneren, althans deze herinneringen toegankelijk te maken voor introspectie, is een verschijnsel dat in het bijzonder intrigeert (zie onder andere Loewenstein, 1993; Vermetten & Bremner, in druk). Dergelijke amnestische fenomenen, waartoe ook het voorkomen van het ‘vergeten’ van de eigen identiteit bij de dissociatieve identiteitsstoornis gerekend kan worden, doen vermoeden dat dissociatie samenhangt met gestoorde geheugenfuncties. Het is inmiddels aannemelijk geworden dat traumatische ervaringen voorboden kunnen zijn van dissociatieve fenomenen, en dat fysieke mishandeling alsook seksueel en emotioneel misbruik bij jonge kinderen kwetsbaarheid voor het ontstaan van dissociatieve stoornissen oplevert (Putnam, 1985; Chu & Dill, 1990; Briere & Conte, 1993; Boon & Draijer, 1993; Coons, 1994; Bremner et al., 1995a). Belangrijk is te vermelden dat niet het trauma, maar de dissociatie als reactie op het trauma de specifieke psychopathologie kenmerkt (Bremner, 1992, 1993; Irwin, 1994; Marmar et al., 1994; Bremner & Brett, 1997a).
Het concept ‘dissociatie’ kreeg, na de herontdekking van het werk van Pierre Janet, opnieuw waardering bij de bestudering van de relatie tussen trauma en geheugen (Nemiah, 1989, Putnam, 1989a en 1998b, Van der Kolk & Van der Hart,1989); op het Freudiaanse concept ‘verdringing’ werd steeds meer kritiek gehoord. Felle discussies over het onderscheid tussen dissociatie en verdringing volgden (zie onder andere Scheflin & Brown, 1996). Dissociatie werd alsnog een gerespecteerd concept; in de classificatie van de DSM-IV werd het woord psychogeen zelfs vervangen door dissociatief (APA, 1994). In vele therapiekamers werd getracht patiënten te leren hoe zij controle over dissociatieve fenomenen zouden kunnen krijgen; ook werd getracht amnestische barrières op te heffen. Bij patiënten kwamen, al of niet met behulp van hypnose, traumata van lang geleden aan het licht.
De discussie over het waarheidsgehalte van traumatische, ‘hervonden’ herinneringen werd een ‘hype’, speciaal de vraag in hoeverre het mogelijk is in therapie pseudo-herinneringen aan te brengen (zie bijvoorbeeld Loftus, 1994; Williams, 1994; Brewin, 1996; Draijer, 1997; Brown et al., 1998).
Sommige auteurs stellen dat seksueel misbruik te ingrijpend is om vergeten te kúnnen worden; een eventueel vergeten van misbruik moet in hun ogen toegeschreven worden aan ‘normaal vergeten’ of aan ‘infantiele amnesie’; in hun ogen maken naïeve therapeuten misbruik van de suggestibiliteit van hun cliënten en patiënten (zie bijvoorbeeld Wagenaar & Crombag, 1995; Merckelbach & Wessel, 1995; Crombag & Merckelbach, 1996).
In ander onderzoek is het argument van de suggestibiliteit van patiënten alsmede de veronderstelling dat herinneringen achteraf in het leven kunnen worden geroepen niet aangetoond of verworpen (Scheflin & Brown, 1996; Bremner et al., 1996; Leavitt, 1997; Brown et al., 1998). Het juridisch aspect van deze oorspronkelijk als wetenschappelijk bedoelde strijd gaat in Amerika gepaard met aanklachten met forse schadeclaims, die ook therapeuten in Nederland angst inboezemen.
Het neurobiologisch onderzoek naar dissociatieve fenomenen en de relatie met trauma en geheugenfuncties is van recente datum.
Er werd aansluiting gezocht bij het neuro-endocrinologisch onderzoek naar de verwerking van stress. Provocatiestudies met onder andere yohimbine en ketamine hebben enige kennis opgeleverd over de wijze waarop het brein met mentale belasting omgaat. Nieuwe, met name de zogenaamde beeldvormende technieken, gaven aanleiding geven tot onderzoeksvoorstellen die men zich tot voor kort niet kon voorstellen. Langs functioneel-anatomisch onderzoek is het nu mogelijk beter inzicht te krijgen in de neurofysiologische en pathofysiologische processen die bij mensen met dissociatieve stoornissen en PTSS van belang zijn. (Het spreekt vanzelf dat het onderzoek met dieren van beperkte waarde is wanneer het om verschijnselen als amnesie gaat.)
Recent geheugenonderzoek over de gevolgen van stress op het geheugen leert dat traumatische herinneringen anders worden gecodeerd en opgeslagen dan herinneringen aan ‘normale’ gebeurtenissen (Cahill et al, 1994; LeDoux 1992, 1996; Southwick et al., 1994; Van der Kolk, 1994; Krystal et al., 1995). In deze bijdrage wordt een overzicht gegeven van de relatie tussen dissociatie en veranderingen in geheugenfuncties bij getraumatiseerde mensen; hierbij worden zowel theoretische als biologische modellen gebruikt.
Geheugenopslag is een variabel proces
Ons geheugen omvat vanzelfsprekend slechts een bescheiden selectie uit de gebeurtenissen die we hebben meegemaakt. Edelman (1989) spreekt over ‘the remembered present’, dit wil zeggen dat wat men als herinnering ervaart in het heden steeds opnieuw moet worden opgebouwd naar aanleiding van zekere aanleidingen (‘cues’).
Men zal zich niet alle diners die men genuttigd heeft kunnen herinneren, maar sommige mogelijk wel. De inhoud van het geheugen wordt dus ook bepaald door de waarde of betekenis (‘value’, Edelman, 1989) die aan een ervaring wordt toegekend. De ervaring leert dat gebeurtenissen met een aanzienlijke emotionele waarde het best herinnerd worden. De meeste ervaringen worden vergeten, of op zijn best slecht onthouden. Inmiddels wordt vrij algemeen aangenomen dat het geheugen niet als een videorecorder werkt en dat ook de term ‘fotografisch geheugen’ een metafoor is – en dus zo goed als niets met fotografie of het geheugen te maken heeft.
Behalve dat de opslag van herinneringen aan gebeurtenissen afhankelijk is van hun emotionele betekenis, geldt dat men zich gemakkelijker herinnert in omstandigheden die verwant zijn aan de omstandigheden waarin de informatie werd opgebouwd. Dit wordt omschreven als ‘context-dependent memory’ (Godden & Baddely, 1975), ‘cue-dependent memory’ (Eich, 1977, 1980) en ‘state-dependent memory’ (LeDoux, 1996).
Gebeurtenissen die verrassend of schokkend zijn (Brown & Kulik, 1977), kunnen ook leiden tot ‘flashbulb memories’, zeer gedetailleerde herinneringen, hypermnesie, voor persoonlijke gegevens in verband met zo’n gebeurtenis. Men weet bijvoorbeeld welke kleren men droeg toen de gebeurtenis plaatsvond en wat men precies aan het doen was. Het geheugen zal beter zijn na persoonlijk belangrijke gebeurtenissen dan na gebeurtenissen van meer algemeen belang. Dit ‘flashbulb’-patroon is kenmerkend voor herinneringen aan enkelvoudige trauma’s. Terr et al. (1996) onderzochten de herinneringen van kinderen die de explosie van het ruimteschip Challenger in 1986 gezien hadden. Bij kinderen die op de tv gezien hadden dat dit ruimteschip explodeerde waren de herinneringen bij de meerderheid consistent en gedetailleerd, ook met betrekking tot de persoonlijke aspecten die ten tijde van de ramp een rol speelden. Kinderen die minder direct bij de ramp betrokken waren, brachten minder detaillering aan, waren minder consistent in hun rapportage en hadden een slechtere ordening van de volgorde van de gebeurtenissen.
Ondanks sterke emotionele betrokkenheid kan de herinnering voor specifieke fragmenten ook niet voor herinnering toegankelijk zijn. Een voorbeeld:
Twee mannen en twee vrouwen zitten op een bank buiten bij een restaurant. Een van vrouwen ziet drie mannen naar hen toekomen. Zij neemt een gevoel van vijandigheid bij hen waar, en ook dat deze gericht is tegen de mannen in haar gezelschap. Dit herinnert zij zich later allemaal goed. Het volgende dat zij zich kan herinneren is dat zij geknield naast haar man zit, die met messteken omgebracht is. Zij was niet onder invloed van alcohol of drugs. Twee jaar later kon zij zich nog niet herinneren dat zij de aanvallers probeerde te beletten naderbij te komen en dat zij in een poging hen af te weren op de grond was gegooid. Ook het feit dat een mes op haar keel was gezet en dat zij zag hoe haar man werd omgebracht kon ze zich niet herinneren. Deze gang van zaken werd gerapporteerd door een getuige die de gebeurtenis gezien had maar niet durfde te interveniëren (Christianson & Engelberg, 1997).
De amnesie van de vrouw in het voorbeeld trad op als reactie op een bijzonder emotionele ervaring. Dit is een typisch dissociatief patroon; hoewel hierin variaties voorkomen treedt op: het onvermogen om zich gebeurtenissen van voor (retrograde amnesie) en van na het trauma (anterograde amnesie) te herinneren, het trauma incluis.
Experimenten in het laboratorium hebben echter aangetoond dat herinneringen kunnen vertroebelen. Hierbij bleek dat de betrouwbaarheid niet altijd zo groot is als in de ‘flashbulb’-hypothese aanvankelijk gedacht werd. Er lijkt een verwantschap te bestaan met wat in de literatuur over hypnotiseerbaarheid beschreven is. Hoog hypnotiseerbare mensen en ‘fantasy prone personalities’ zijn suggestibel en hebben in het algemeen moeite met het in de gaten houden van de grens tussen werkelijkheid en fantasie (Lynn & Rhue, 1986, 1988). Ondanks de complexiteit van de variabelen in dergelijk onderzoek zijn er aanwijzingen dat hoog-hypnotiseerbare personen eerder verkeerde informatie in het geheugen opslaan dan laag-hypnotiseerbare mensen (Labelle et al., 1990; Barnier & McConkey, 1992). Hoewel nog gezocht wordt naar een betrouwbare manier om het scheppen van pseudo-herinneringen in therapieën vast te stellen (Brown, 1995), zijn er voldoende aanwijzingen om aan te nemen dat hypermnestische suggesties onjuiste veronderstellingen in de hand werken en zowel het ‘geloof’ in waar als in niet-waar versterken (Erdelyi, 1996; Kihlstrom, 1997). Om deze reden is het verstandig in een forensische of therapeutische situatie af te zien van hypermnestische procedures om het geheugen te verbeteren of te versterken; dit standpunt is analoog aan dat van de American Society of Clinical Hypnosis (ASCH) (Hammond et al., 1995).
Dissociatieve processen gaan gepaard met een bijzonder type geheugenopslag
Dissociatieve of psychogene amnesie wordt gekenmerkt door ‘gaten’ in de herinnering, die minuten tot dagen in beslag nemen. Toegang tot de traumatische herinneringen treedt vaker op wanneer de patiënt zich in een gedissocieerde toestand bevindt met symptomen als derealisatie en depersonalisatie. Gefragmenteerde of afgesplitste geheugeninhouden kunnen de aandacht wegtrekken van de actuele werkelijkheid. Dissociatie gaat ook gepaard met de betrekkelijke afwezigheid van de integratieve functies van identiteit, geheugen en bewustzijn. Het is een toestand van het bewustzijn die vaak ‘state dependent’ blijkt. Het is gebleken dat dissociatieve symptomatologie vaker voorkomt na een trauma en dat het optreden van dergelijke symptomen posttraumatische stress-klachten kan voorspellen. Na de ramp met de Estonia-veerboot in 1995 traden bij iets minder dan de helft van de overlevenden dissociatieve symptomen op. Zij rapporteerden de genoemde gevoelens van derealisatie en depersonalisatie en een veranderd tijdsbesef; deze dissociatieve symptomen bleken betrouwbare voorspellers van posttraumatische reacties (Eriksson & Lundin, 1996). De herinnering aan het trauma trad in gedissocieerde vorm op. Deze eigenaardigheden doen vermoeden dat dergelijke herinneringen op een bijzondere manier worden opgeslagen en opgehaald. De korte-termijn-verstoringen hangen wellicht samen met een ongebruikelijk verhoogde aandachtsfocus (‘tunnel memory’) en aan dissociatieve reacties tijdens de traumatische gebeurtenissen. Een vergelijking biedt de posthypnotische amnesie die met een klassieke hypnotiseerbaarheidsschaal als de Stanford Scale of Hypnotic Susceptibility bij hoog-hypnotiseerbare personen kan worden bereikt: tijdens de hypnose wordt het subject gesuggereerd een opdracht uit te voeren (bijvoorbeeld de schoenzolen aanraken) zonder dat deze weet met welke aanwijzing (bijvoorbeeld tikken op de tafel) de opdracht wordt gegeven. Het opheffen van de amnesie door de aanwijzing te onthullen roept vaak verwarring op, waarbij dan ook nog niet altijd de herinnering aan de aanwijzing toegankelijk wordt.
Dissociatieve amnesie kan jaren voortduren. In de inmiddels veel geciteerde studie van Williams bleek 38% van een groep vrouwen die als kind werd misbruikt zich dit jaren nadien niet te herinneren (Williams, 1994b; 1995). Er is een leeftijds- en ‘dosis’-relatie met het optreden van significante amnesie. Hoe jonger op het moment van het trauma, hoe groter de mogelijkheid van het optreden van amnesie (Briere & Conte, 1993); de verklaring moet dan vermoedelijk ‘infantiele amnesie’ zijn. Daarnaast doet amnesie zich ook vaker voor wanneer het misbruik zich over langere tijd uitstrekte (Terr, 1991; Herman, 1992; Ross et al., 1991).
De dissociatieve identiteitsstoornis is een bijzondere manifestatie van dissociatieve symptomen in het algemeen en dissociatieve amnesie in het bijzonder. Het gaat om verschillende collecties autobiografische herinneringen die als (min of meer volledig) separate entiteiten ‘voortleven’. Verondersteld wordt dat deze discontinuïteiten samenhangen met een veranderde activiteit in de hersengebieden die betrokken zijn bij integratieve processen. Hiernaar vindt met behulp van Positron Emission Tomografie (PET) in Nederland aan de Rijksuniversiteit Groningen onderzoek plaats (Korf & Nijenhuis, persoonlijke mededeling, 1998).
Posttraumatische reacties zijn dissociatieve reacties op traumatische ervaringen
Patiënten met een posttraumatische stress-stoornis (PTSS) hebben uiteenlopende manifestaties van veranderingen in het normale geheugen, inclusief deficiënties in de functies van het expliciete en impliciete geheugen. Er bestaat een ruime ‘overlap’ tussen PTSS en de dissociatieve stoornissen. In een studie uit 1992 werd aangetoond dat uit een populatie oorlogsveteranen degenen met de diagnose PTSS een significante hogere score op de DES lieten zien dan veteranen die niet aan de criteria voor PTSS voldeden. Het verschil bleef gehandhaafd na statistische correctie voor de mate van blootstelling aan trauma (Bremner et al., 1992). Overeenkomstig Janets gedachte dat dissociatie als gevolg van een extreem trauma het risico op dissociatieve reacties op volgende traumata vergroot, lieten de oorlogsveteranen met PTSS een toename van dissociatieve reacties zien op belastende gebeurtenissen na hun diensttijd. Zij hadden ook toegenomen dissociatieve reacties tijdens het terughalen van de traumatische gebeurtenissen. Dissociatieve reacties hangen samen met hypnotiseerbaarheid: patiënten met PTSS vertonen een hogere hypnotiseerbaarheid dan personen met een vergelijkbare blootstelling aan trauma zonder symptomen van PTSS (Spiegel et al, 1988). Ook de leeftijd lijkt een belangrijke variabele te zijn. Mensen die tijdens hun kinderjaren blootgesteld werden aan misbruik vertonen meer dissociatieve symptomatologie dan volwassenen die dit misbruik bespaard bleef (Putnam et al., 1985; Chu & Dill, 1990). Marmar et al. vestigden de aandacht op peritraumatische dissociatie die een voorspellende waarde had voor het later optreden van PTSS (Marmar et al., 1994, 1996). Carlier et al. (1996) beschreven dat symptomen van PTSS op hun beurt een voorspellende waarde hebben voor het optreden van dissociatieve symptomen.
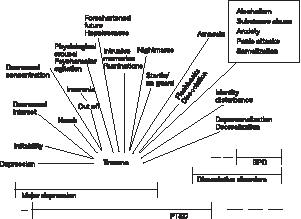
Traumatische stress komt terug in verschillende psychopathologische categorieën (zie figuur 1). Verschillende symptomen van depressie, PTSS, dissociatieve stoornissen en de borderline persoonlijkheidsstoornis vertonen verband met traumatische ervaringen. De dsm-iv (apa, 1994) draagt niet bij tot een beter begrip van de wijze waarop trauma tot psychopathologie kan leiden. Dissociatieve stoornissen zijn losgekoppeld van PTSS, terwijl onderzoek robuust aantoont dat beide tegelijk voorkomen. Van de patiënten met een borderline persoonlijkheidsstoornis kent 80% een geschiedenis van vroeg seksueel misbruik (Herman, 1992), evenals 80% van de patiënten met een dissociatieve identiteitsstoornis (Putnam, 1991). Een deel van de symptomen van de diverse soorten depressie zijn gelijk aan die van PTSS (Bremner et al., 1998).
Neurobiologische codering bij geheugenopslag in afhankelijkheid van emotie
Al in het begin van deze eeuw was duidelijk dat emotie enerzijds het geheugen kon versterken en hypermnesie bevorderen, maar dat emotie ook hypomnesie kon doen ontstaan. Hierbij bleek ook dat endogene stresshormonen in een omgekeerde U-vormige relatie stonden tot het geheugen waarbij een bepaalde dosis versterkend kon werken, en een toegenomen dosis juist verminderend (Stratton, 1919). Pas veel later kwam uit zowel cognitief-psychologisch als psychofysiologisch onderzoek de hypothese naar voren dat er verschillende neurobiologische systemen betrokken zijn bij het opslaan van emotionele ervaringen, en dat bij het opslaan van emotionele ervaringen andere geheugensystemen betrokken zijn dan bij niet-emotionele ervaringen. Verschillende auteurs hebben overigens tegen dit onderscheid geargumenteerd (Neisser, 1982; McCloskey et al., 1988). Toch blijkt het geheugen voor emotionele gebeurtenissen meer accuraat en minder gevoelig voor verandering in de tijd dan gebeurtenissen die geen emotionele reactie teweegbrengen en vrijwel puur aan aandachtsvariabelen onderhevig zijn (Christianson & Loftus, 1987; Conway et al., 1994; Cahill & McGaugh, 1995).
Twee neurobiologische geheugenopslagsystemen blijken in het bijzonder betrokken te zijn tijdens en vlak na traumatische stress: (1) endogene stresshormonen, in het bijzonder de catecholaminen noradrenaline en adrenaline, en de glucocorticoiden met cortisol als stresshormoon, en (2) de amygdala in nauwe relatie tot de hippocampus. In niet-emotionele situaties zijn deze systemen relatief inactief. Zij worden verhoogd actief bij emotionele en stressvolle leersituaties en ‘hebben’ dan als doel om de sterkte van een geheugenspoor, of de sterkte van de leerervaring proportioneel te laten zijn aan het belang van de ervaring voor het organisme (Cahill, 1997; McGaugh & Cahill, 1997).
Het ‘normale’ geheugen versus het geheugen bij trauma
Om een idee te verkrijgen over de met stress samenhangende veranderingen in het geheugen, volgt eerst een korte beschrijving van het ‘normale’ geheugen. Hoewel de literatuur over de neurobiologie van het geheugen enorm uitgebreid is, kunnen gelukkig enkele principes onderscheiden worden:
- sporen van geheugeninhouden zijn gelokaliseerd in verschillende en meerdere regionen in het brein;
- verschillende vormen van geheugen corresponderen met verschillende neuronale circuits (Kandel & Schwartz, 1985);
- ervaringsgewijs opgebouwde synaptische plasticiteit als vorm van biologisch geheugen is een wijze waarop instinctieve processen zich aanpassen aan veranderende levensomstandigheden;
- leren, het geheugen en geheugensporen (engrammen) verwijzen steeds naar ervaringsafhankelijke plastische veranderingen in het brein (Vanderwolf & Cain, 1994).
- het geheugen integreert perceptieve ervaringen (olfactorisch, sensorisch, visueel, kinestetisch, akoestisch);
- het geheugen is ononderbroken, en vermindert langzaam aan accuratesse in de tijd;
- het geheugen is meestal ‘declaratief’, het kan uitgedrukt worden in woorden en symbolen (Van der Kolk et al., 1996).
In hedendaagse indelingen van het geheugen wordt het oude onderscheid in episodisch, semantisch en procedureel geheugen grotendeels verlaten. De indeling die tegenwoordig gebruikt wordt (figuur 2), is: (1) expliciet/declaratief geheugen (bewuste herinneringen aan [unieke] feiten inclusief werkgeheugen) en (2) impliciet/nondeclaratief geheugen (Squire & Zola-Morgan, 1991; Schacter, 1995).
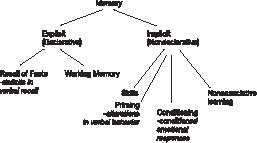
In deze indeling wordt de inhoud van het impliciete geheugen gedefinieerd als herinneringen die niet toegankelijk zijn voor introspectie; het expliciete geheugen is wel toegankelijk voor het bewustzijn. Het impliciete geheugen heeft als taken: ‘priming’, conditionering en niet-associatief leren. Om zich bij ‘priming’ iets voor te stellen kan men zich een experiment indenken waarbij een deel van een woord wordt aangeboden en ons gevraagd wordt het eerste woord te noemen dat wij ons herinneren. Conditionering is substitueren van een stimulus. Voorbeeld: een lichtflits of een geluidsprikkel kan als aanvankelijk (neutrale) ongeconditioneerde stimulus na enige tijd een consistente fysiologische en emotionele respons doen optreden. Een neutrale stimulus heeft na het leren een voorspellende waarde voor de reactie van het organisme gekregen en is vervolgens dus niet meer neutraal. Volgens dit paradigma van impliciete geheugenfuncties is het leren ingebed in een geconditioneerde respons, zonder dat er interferentie met het bewustzijn is.
In het algemeen wordt onder geheugenopslag verstaan: codering (encoding), opslag of consolidatie (storage), en het terugvinden van informatie (retrieval). De codering is de eerste neerslag van een geheugenspoor of -element. Opslag betreft het bewaren van het geheugenspoor over langere tijd. Het verwante begrip consolidatie verwijst speciaal naar het proces dat plaatsvindt als het geheugenspoor wordt opgeslagen, wat, theoretisch, de periode is waarin het spoor of de inhoud te wijzigen of te vervormen is. Bij het terughalen van informatie is belangrijk dat informatie over verschillende corticale gebieden coherent word samengevoegd tot een volledige herinnering, bestaande uit verschillende modaliteiten (beeld, geluid, etc.), in relatie tot ruimte en tijd.
Posttraumatische geheugenveranderingen maken het moeilijk de bestaande theorieën over de geheugenfuncties te handhaven. Herinneringen aan trauma hebben bepaalde kenmerken. In de volgende paragrafen vormen deze kenmerken het motief voor een neurobiologische modelvorming.
Tijdens trauma
Aandacht: er is sterke aandacht voor in het oog springende informatie, er is een verminderd aandachtsfocus op contextuele informatie.
Perceptie: de perceptie wordt vervormd door dissociatieve processen tijdens het trauma.
Opslag: met name non-verbale geheugenopslag, dus visuele beelden en andere zintuiglijke informatie, met emotionele elementen van het trauma, worden verdeeld over het brein, maar ze worden geassocieerd opgeslagen, zij vormen het motief voor patronen van herhaling en herinnering.
Markante informatie wordt opgeslagen in het lange-termijngeheugen, de gecodeerde informatie reflecteert een verlies van contextuele informatie door dissociatieve processen en veranderde aandachtsvariabelen.
Na trauma
Aandacht: stimulusgeneralisatie; de aandachtsfuncties worden gestuurd door het trauma.
Herhaling: herinneringen worden gevormd in een doorlopend proces; de inhoud en de associaties kunnen variëren in de tijd.
Herinnering: bewuste toegang kan verdwenen of verminderd zijn (dissociatieve amnesie).
Intrusieve beleving van de herinnering; na een eerste herinnering gaat het herinneren almaar door.
Onbewuste mechanismen van het zich herinneren zijn prominent, de herinnering bevat exteroceptieve en interoceptieve (emotionele en fysiologische) aanleidingen.
Bewustzijnstoestanden die samenhangen met een verminderd niveau van alertheid (slaap, hypnose) en/of een vermindering van executieve functies (bijvoorbeeld alcohol-intoxicatie) kunnen de weerstand tegen de herinnering verminderen.
Sommige herinneringen hangen samen met een verlies van perspectief en worden eerder herbeleefd dan herinnerd.
Eenmaal herinnerd zijn de herinneringen slechts toegankelijk op een relatief inflexibele manier, als weerspiegeling van verstoringen in strategische herinneringsprocessen.
Het herinneren kan gepaard gaan met intense emotionele reacties en dissociatie.
Veel discrete kwaliteiten van de traumatische ervaring verminderen niet in de loop van de tijd.
Neurale informatieverwerkende systemen als verklarend paradigma voor disociatieve processen
Tegenwoordig vormen ‘connectionistische modellen’, ‘parallel verdeelde systemen’ en ‘associatieve neurale netwerken’ het opkomend paradigma voor de vertaling van informatieoverdracht in het brein (Hopfield, 1982; Parks et al., 1991; Jeffrey et al., 1997). Op deze gebieden kunnen neurowetenschappen en cognitieve psychologie elkaar aanvullen. Verondersteld wordt dat neurale netwerken interconnectieve systemen vormen en zich onderling verbinden. Taal, cognities, motorische activiteit en affecten zijn als een soort ‘knopen’ in deze systemen opgeslagen. In het brein bestaan ‘knopen’ uit subcorticale structuren waaronder hippocampus, amygdala, hypothalamus en het reticulaire activerende systeem, die via neuronale verbindingen, synapsen en transmitters in nauw contact met elkaar staan. Maar ook neocorticale structuren staan hiermee in verbinding, zodat domeinen en vervolgens netwerken ontstaan. De onderlinge verbindingen van neurale structuren maakt functies mogelijk die niet verklaard kunnen worden uit de afzonderlijke structuren.
Voorbeeld: een wandelaar loopt op een mooie zomerdag in de bergen. Hij ziet een slang. Hij merkt dat zijn hartslag stijgt en zijn ademhaling versnelt. Wat gebeurt er sequentieel? Het vaststellen dat een slang in de bergen is waargenomen vindt plaats in een parallel verdeeld neuronaal systeem: visuele informatie wordt gevoegd bij motorische, en het beeld van de slang wordt vergeleken met reeds opgeslagen beelden van slangen. Deze slang lijkt op die uit de dierentuin. Komen hier slangen voor? Heeft de slang het op mij gemunt? Waar stel je een slang tevreden mee? Hoe houd ik de slang voor de gek en kom ik hier weg? De informatieverwerking vindt in milliseconden plaats via activatie van diverse neurale units, die op biochemisch niveau gereguleerd worden door het vrijkomen van neuromodulatoren; input wordt steeds vergeleken met bestaande informatie op basis waarvan onbewuste stuurprocessen optreden in het brein waarna uiteindelijk de ‘final pathway’ en (bewust) gedrag optreedt. Bij het ‘interconnecteren’ van netwerken om geheugenprocessen mogelijk te maken zijn twee specifieke eigenschappen van belang: het geheugen moet generiek zijn (bijvoorbeeld het herkennen van slangen als diersoort uit de dierentuin) maar moet tegelijkertijd specifiek zijn (deze slang is niet een slang uit de dierentuin).
Geheugensporen worden gevormd voor voorwerpen, geluiden, of voor andere modaliteiten, steeds in een voortdurende plaats- en tijdverankering, afhankelijk van de aandacht. De neuromodulatoren die vrijkomen als reactie op de ervaring kunnen een meer of minder versterkend effect op de ervaring zelf hebben, afhankelijk van onder andere de hoeveelheid en van het soort neuromodulatoren dat vrijkomt. De ervaring zelf bepaalt aldus of een geheugenspoor sterk of zwak zal zijn.
De toegankelijkheid van de ervaring kan afhangen van de organisatie van de opslag van het geheugenspoor en laat zich beschrijven in het model van parallelle verdeelde systemen (‘parallel distributed processing’, PDP) (Rumelhart & McLelland., 1986). Dit is een gebiologiseerd model dat begrijpelijk tracht te maken hoe traumatische ervaringen als sensorische input afgesplitst kunnen worden, gedissocieerd van het bewuste geheugen. Het model beschrijft dat er verschillende routes mogelijk zijn waarlangs ervaringen geïntegreerd kunnen worden en kunnen corresponderen met verschillende geheugensystemen. Bestaande connecties in informatieverwerkende systemen kunnen de weg beïnvloeden waarlangs patronen voor toekomstige verwerking plaatshebben. De sterkte van de onderlinge connecties kan in belangrijke mate bijdragen tot de sterkte van het geheugenspoor, de route waarlangs, en de mate waarin het geheugenspoor geïntegreerd is met andere geheugensystemen. De niet toegankelijke herinnering kan toegankelijk worden als een sterk aspect van een gebeurtenis opnieuw voorkomt ofwel als sensorische of affectieve input, waardoor sommige units die eerder hebben meegedaan bij de representatie van die episode, geactiveerd worden. De activering kan vervolgens andere units activeren en een cascade van consequente reacties doen ontstaan. Dit model geeft echter ook aan hoe complex verweven en ook verward het geheugen kan zijn. Het is dan ook niet zeer verwonderlijk dat aspecten van het functioneren van het geheugen vatbaar kunnen zijn voor een bepaalde mate van vervorming of soortgelijke problemen (Edelman, 1989).
Zicht op traumatische stress: een voortzetting van het werk van Selye
Traumatische stress doet een appèl op verschillende biochemische processen. De intensiteit van de respons en de veranderingen die deze teweegbrengt kunnen tot enig onderscheid leiden tussen stress en traumatische stress. Voor de stressrespons is het werk van Selye (1936) elementair. Een basale veronderstelling van Selye is dat zowel fysieke als emotionele stress centrale en perifere processen in gang zet die ervoor dienen om de homeostase te bewaren. Hierbij kan verhoogde hartslag ontstaan, bloeddrukstijging, versnelde ademfrequentie gepaard aan verhoogde alertheid en waakzaamheid. Tegelijkertijd is er een verminderde opname en vertering van voedsel, de darmmotiliteit neemt af, er is een verminderde reproductieve functie en een verzwakte immuunrespons. Bij Selye hield het in kaart brengen van de perifere respons goeddeels op. Waar hij ophield gingen wetenschappers als Sapolski, McEwen, LeDoux en Cahill verder met hun pogingen de stressrespons in kaart te brengen, ook bij centrale processen in het brein.
Belangrijk uitgangspunt bij het onderzoek naar invloed van stress op het brein in het algemeen en op geheugenfuncties in het bijzonder is dat individuen die worden blootgesteld aan traumatische stress dezelfde neurobiologische mechanismen gebruiken als individuen die aan normale stress blootstaan. Het onvermogen om adequaat om te gaan met traumatische stress kan leiden tot ernstige neurobiologische veranderingen en consequenties. Iedere neurobiologische aansturing wordt verondersteld onwillekeurig te zijn. Leidt een succesvolle aanpassing tot herstel van een normale homeostatische balans, een mislukte adaptatie kan een evenwicht doen ontstaan dat, hoewel het eveneens stabiel is, sterk afwijkt van een normale neurobiologische standaard. Deze abnormale standaard wordt allostase genoemd (McEwen, 1995).
De fysiologische veranderingen die optreden als gevolg van stress maken het ons mogelijk om op tijd de trein te halen, een vergadering te leiden, maar ook om ’s morgens op te staan en uit bed te komen. Stress stelt ons in staat extra bronnen te activeren die in de actuele situatie als noodzakelijk geïnterpreteerd worden. In de acute situatie kan de reactie nuttig zijn, maar de arousal mag niet blijven voortbestaan. Immers, in een blijvend verhoogde arousal kan een toegenomen ‘allostatic load’ aanleiding zijn voor ingrijpende veranderingen in neurobiologische systemen en het gedrag, met psychopathologie als gevolg (Mazure, 1994; McEwen & Schmeck, 1994; Friedman et al., 1995). Waar in eerste instantie een reactie op stress adaptief kan zijn en het organisme in staat stelt te overleven kan, wanneer deze toestand niet overgaat, het een aanleiding zijn voor het ontstaan van ernstige ziekte: ‘What is in the short term adaptive, may in the long run result in pathology’ (Bremner et al., 1993).
Centrale neurobiologische en neuro-anatomische systemen die betrokken zijn bij reacties op traumatische stress
Neurobiologische opslagmechanismen
Opslag van informatie in het lange-termijngeheugen (‘long term memory’, LTM) wordt gemoduleerd door een endogeen neurobiologisch systeem. Dit systeem is gewoonlijk inactief en niet emotioneel geëngageerd, maar kan actief worden in belastende situaties waarbij de sterkte van het geheugenspoor proportioneel is met het belang of de betekenis voor het individu. Zoals al eerder gezegd zijn twee neurobiologische elementen, of systemen fundamenteel: (1) endogene stresshormonen, in het bijzonder de catecholaminen noradrenaline en adrenaline, en (2) als neuro-anatomische basis van het geheugen: de amygdala en hippocampus.
Endogene stresshormonen
Ad 1. Catecholaminen
Als iemand na een leerervaring een injectie met adrenaline, een stof die op de b-adrenerge neuroreceptor werkt, krijgt toegediend neemt het geheugen voor die leerervaring toe (Gold, 1977; Cahill et al., 1994). Een lage dosis verbetert het geheugen, bij een hoge dosis functioneert het geheugen slechter. Cahill onderzocht het effect van een b-blokker en vond dat propanolol, een b-adrenerge blokker, een verminderde geheugenfunctie veroorzaakte. Propanolol werd toegediend kort voor een emotionele situatie (namelijk het vertellen van een emotioneel geladen verhaal). Een groep die een placebo kreeg toegediend liet na een tijdsinterval van een week, een toegenomen geheugen zien voor het verhaal; de propanolol-groep toonde een verminderd geheugen. Op de herinnering aan een neutraal verhaal had propanololtoediening geen aanwijsbare uitwerking. Geconcludeerd werd dat neurobiologische aangrijpingspunten voor geheugenprocessen aanwezig zijn en dat de b-adrenerge receptoren een rol spelen bij de modulering van geheugensporen (Cahill et al., 1994).
Wat is nu de rol van deze receptoren? De b-adrenerge receptoren van afferenten van de parasympaticus (n. vagus) projecteren naar de kern van de tractus solitarius in het ruggenmerg, waardoor de locus coeruleus gestimuleerd wordt om adrenaline in de amygdala (en hippocampus) te stimuleren (Packard et al., 1995; LeDoux, 1996). Deze receptoren met hun aangrijpingspunt op centrale limbische structuren dragen bij aan de modulering van impliciete emotionele herinneringen en expliciete herinneringen aan emotie.
Men probeerde de adrenerge respons nog specifieker te moduleren. Door provocatiestudies met yohimbine bewerkstelligde men bij patiënten met PTSS een toegenomen noradrenaline-release in het brein. Yohimbine is een alfa-2-adrenerge receptor-antagonist die zorgt dat er presynaptisch meer noradrenerge activiteit plaatsvindt. Gaf men dit aan PTSS-patiënten, dan zag men het optreden van flashbacks, en er ontstond zeer hevige angst. Bij controlepersonen bleef deze reactie afwezig; evenmin trad zij op bij mensen met een paniekstoornis. Het metabolisme gemeten met PET ([18F]fluoro2deoxyglucose, FDG) liet zien dat PTSS patiënten een verminderd metabolisme hadden in neocorticale structuren (orbitofrontaal, temporaal, parietaal en prefrontaal). De controle-proefpersonen lieten daarentegen een toegenomen metabolisme in deze regionen zien. De PTSS-groep had een verlaagd metabolisme in de hippocampus; dit werd niet in de controlegroep gezien. Deze bevindingen lijken consistent met de hypothese dat er een toegenomen noradrenaline-release is in het brein na toediening van yohimbine (Southwick et al., 1991; Southwick et al., 1993). De PTSS-patiënten blijken dus overmatig sensitief voor yohimbine, zij maken meer noradrenaline vrij en laten een verminderd metabolisme van het brein zien. Er ontstaat een verhoogde selectieve attentie of een verhoogde neuronale activiteit voor relevante stimuli en een vermindering voor wat er op de achtergrond gebeurt. Gegeven de rol van noradrenaline als neuromodulator bij encodering van geheugeninhouden en bij herinnering, suggereert dit gegeven dat verhoogde release in de hippocampus onder stress samenhangt met een pathologisch type herinnering dat kenmerkend is voor de herbelevingen die zich bij PTSS patiënten voordoen. De chronische hyperarousal en vigilantie lijkt te passen bij de verhoogde gevoeligheid van het brein voor catecholaminen (Southwick et al., 1994).
Dezelfde bevindingen werden verkregen na toediening van ketamine, een niet-competitieve antagonist van de N-methyl-D-aspartase (NMDA)-receptor, een receptortype dat veel voorkomt in de hippocampus en sterk betrokken is bij de opslag naar het lange-termijngeheugen (Long Term Memory, LTM) (Krystal et al., 1995).
Ad 2. Glucocorticoïden
Het betreft hier een ander biochemisch neuro-endocrien systeem dat een belangrijke rol speelt bij de coördinatie van het lichaam bij stress. Het wordt gevormd door de as van de hypothalamus, de hypofyse en de bijnier (HPA-as). Net als bij de catecholaminen is er ook hier sprake van een omgekeerde U-vormige relatie bij de modulering van geheugeninformatie. Hormonale stimulering door neuropeptiden in het brein stimuleren de release van de corticotropine releasing hormoon (CRH) in de hypothalamus, te zamen met enkele andere stoffen, waaronder vasopressine, waardoor in de hypofyse een stof wordt afgescheiden, adrenocorticotroop hormoon (ACTH), dat de bijnieren aanzet tot het vormen van adrenaline en cortisol. CRH wordt gedistribueerd onder andere over amygdala, hippocampus, prefrontale schors en de gyrus cinguli, locus coeruleus, thalamus, het periaquaductale grijs en cerebellum.
De stressrespons wordt in de literatuur traditioneel gedefinieerd door de hoogte van het geproduceerd cortisol. De belangrijkste functie van cortisol is gelegen in het tijdig afbreken van de biologische stressrespons. De biologische reacties verminderen door onderdrukking van de HPA-as via een negatieve feedback van cortisol op onder andere de hypofyse en hypothalamus.
Hoge cortisolspiegels worden traditioneel geassocieerd met stress, en wel zo dat een stressreactie wordt afgemeten aan de hoeveelheid geproduceerde cortisol (Chrousos & Gold, 1992). Het is bekend dat bij depressieve stoornissen ook een verhoogd cortisol wordt gevonden, hetgeen al een twintigtal jaren geleden aanleiding was om de rol van negatieve levenservaringen en stress te bestuderen bij de etiologie van depressie. Verschillende onderzoeken lieten in de 24-uurs urine van PTSS-patiënten een paradoxale lagere hoeveelheid cortisol zien dan verwacht (Mason et al., 1986; Pitman & Orr, 1990; Yehuda et al., 1995a). Cortisol bleek geen weerspiegeling te zijn van de blootstelling aan trauma, maar wel gerelateerd aan de aanwezigheid van symptomen. Men onderzocht cortisolreceptoren waarbij bleek dat de sensitiviteit (stressgevoeligheid) was toegenomen door toename van het aantal receptoren (Yehuda et al., 1995b). Vanwege het feit dat de glucocorticoid-receptoren de sterkte van de negatieve feedback bepalen, werd gedacht dat een toename in receptoractiviteit een verlaging van de cortisolspiegel met zich meebrengt bij patiënten met PTSS. Het is echter ook bekend dat lage hormoonspiegels gepaard gaan met een toename van receptoren.
(Nor)adrenaline en glucocorticoiden doen globaal hetzelfde, maar op een verschillende tijdschaal. Adrenaline heeft een ander aangrijpingspunt omdat het de bloed-hersenbarrière niet passeren kan, het geeft informatie via de nervus vagus naar de locus coeruleus, die te beschouwen is als de alarmbel van het centrale zenuwstelsel. Vandaar gaat informatie naar de amygdala en de hippocampus. Met betrekking tot het onderscheid tussen adrenaline en cortisol gebruikt Sapolski (1996) de metafoor van een land dat door een agressor wordt aangevallen. In eerste instantie haalt de verdediging de geweren uit de opslagplaatsen en brengt het bestaande leger in stelling. In tweede instantie, op iets langere termijn, roept het reservisten op en schakelt het fabrieken in en gaat over op de productie van wapens en kogels. Adrenaline haalt de wapens uit de opslagplaatsen en cortisol past het productieproces aan. Op celniveau veranderen adrenaline en noradrenaline de functie van enzymen die al in de cel aanwezig zijn. Glucocorticoïden (cortisol) werken meer op genniveau en zetten aan tot synthese van meer en van andere enzymen.
Beide systemen reageren niet steeds even sterk op verschillende stressoren; angst en geweld leiden tot een toename van adrenaline, en depressie tot een toename van glucocorticoïden. Een dier dat een gevecht verliest heeft meer last van de effecten van glucocorticoïden: een dier dat zich doodstil houdt in het hoge gras omdat er een leeuw in de buurt is, loopt over van de adrenaline.
De neuro-anatomische basis van het geheugen
Stress resulteert in een wijd verspreid effect op geheugenfuncties en op de hersenregionen die bij het geheugen betrokken zijn. Deze hersengebieden (hippocampus en naburige corticale regionen, prefrontale corticale plaatsen, amygdala, mediale thalamus, partitale cortex, en sensorische neocorticale gebieden) zijn alle betrokken bij blootstelling aan stress. Deze regionen mediëren op hun beurt de stressrespons. Specifieke functies voor het geheugen vinden plaats in hippocampus en amygdala.
Ad 1. De rol van de amygdala
De amygdala, een groep gespecialiseerde hersencellen, vormt een kleine kern, is gepaard aangelegd, en bevindt zich juist voor de hippocampus in het brein. Deze kern speelt een belangrijke rol als mediator van het emotionele geheugen, en wordt vandaar wel omschreven als ‘the emotional brain’ (LeDoux, 1996). De amygdala integreert informatie die nodig is voor de uitvoering van de stressrespons, waarbij emotie wordt geïntegreerd met en geïnitieerd door informatie uit de externe omgeving. Als iemand bij het wandelen in de bergen een slang ziet zal in eerste instantie het signaal van een zich snel bewegende lijn via het thalamische baansysteem de amygdala bereiken. Cellen in de amygdala zullen in een geëxciteerde toestand geraken totdat informatie van de cortex komt en bevestigt dat het om een onschadelijke slang gaat, waarna doelmatige gedragsverandering kan optreden. Als nu een meer gedetailleerde corticale analyse oplevert dat de stimulus slechts een regenworm of, nog erger, een boomtak is, zullen verdere angstresponsen geremd worden. Als dezelfde slang in de dierentuin gezien wordt zal – hoewel de thalamus en de cortex dezelfde informatie signaleren – geen angstrespons optreden omdat de stimulus wordt geassocieerd met een veilige context, zoals wordt geleverd door informatie uit de hippocampus naar de amygdala.
De amygdala heeft met betrekking tot het geheugen drie functies:
- deze kern speelt een belangrijke rol bij angstconditionering,
- is betrokken bij de extinctie van cognitieve associaties naar het oorspronkelijke trauma,
- de amygdala integreert sensorische en cognitieve input van verschillende hersenregionen (Squire, Zola &Morgan, 1991).
Bij de angstconditionering wordt een onschadelijke geconditioneerde stimulus (CS) gepaard aan een aversieve, ongeconditioneerde stimulus (US), waarbij een CS-US-koppeling optreedt. Het effect van stress op de amygdala, hippocampus en prefrontale cortex kan resulteren in context-onafhankelijke krachtige angstresponsen die niet uit te doven zijn. Als de corticale input op de amygdala wegvalt kan elke stimulus die overeenkomsten met deze CS vertoont een respons geven die resulteert in overgeneralisatie van angstresponsen. Men hoeft zich daarbij niet bewust te zijn van bijdrage van de elementaire kenmerken van de angststimulus; men kan evengoed spontane niet geprovoceerde angstresponsen vertonen. Deze verschijnselen vertonen sterke overeenkomst met de symptomen van PTSS. Hierbij kan stress als reactie op bedreigende stimuli dezelfde reacties te zien geven als de anticipatie op bedreigende stimuli.
Veel kennis van de neurale mechanismen waarbij de amygdala is ingeschakeld is afkomstig uit experimenteel onderzoek. Tegenwoordig biedt ook beeldvormend onderzoek de mogelijkheid om hypothesen te toetsen en eventueel aan te scherpen. Bij dierstudies bleek dat de eerste koppeling via het thalamo-amygdala-systeem binnen 15 milliseconden na het aanbieden van bijvoorbeeld een geluidssignaal optreedt. Men heeft ontdekt dat in dit proces plastische veranderingen in de amygdala plaatsvinden. Er vindt een ‘fijntuning’ plaats van neurale receptieve velden in de amygdala die resulteert in een toegenomen representatie op neuronaal niveau van de CS. Als de centrale kern van de amygdala geblokkeerd wordt, verdwijnt een geconditioneerde ‘startle’, terwijl stimulatie ervan leidt tot toename.
Patiënten met laesies in de amygdala lieten een verminderd aanleren van autonome geconditioneerde responsen zien in het angstconditionerings-paradigma, terwijl zij zich note bene bewust waren van de aard van de koppeling, dus zowel van de CS als van de US. Patiënten met een intacte amygdala, maar met uitgebreide hippocampale laesies laten normale geconditioneerde responsen zien, maar missen de declaratieve (bewuste) herinnering van de CS-US-koppeling.
De amygdala krijgt aldus parallelle input van verschillende kanalen. Een verstoorde balans in de input kan tot krachtige ondoelmatige angst leiden, zoals gezien wordt bij PTSS.
Ad 2. De rol van de hippocampus
De hippocampus is een langgerekte structuur, ontstaan als olfactorisch deel van de cortex. De structuur is gepaard aangelegd. In feite is het gemodificeerde cortex van de temporale schors. Ieder type sensorische gewaarwording veroorzaakt specifieke activatie van de hippocampus, variërend van woede tot seksuele activiteit. Elektrische stimulatie kan epileptische aanvallen veroorzaken. Binnen het geheugen is de functie van de hippocampus het opslaan van geheugensporen door deze te categoriseren en er een spatio-temporele code aan toe te voegen.
Dit laatste is erg belangrijk: er kan pas een beeld ontwikkeld worden wanneer voortdurend ruimte en tijd aan elkaar gekoppeld worden en wanneer het beeld van plaats verandert. De integratie van de perceptie verloopt aldus via een ‘temporal succession’ (Edelman, 1989).
De hippocampus is van betekenis bij het declaratieve geheugen, samen met de nabijgelegen neocorticale structuren (entorhinale, perirhinale en parahippocampale cortex) en de dorso-mediale nucleus van de thalamus (Mishkin, 1978, Zola-Morgan et al., 1989; Squire and Zola-Morgan, 1991). De dorsolaterale prefrontale cortex (sulcus principus of mediale frontale gyrus) is hierbij betrokken wanneer taken uit het werkgeheugen worden gehaald (Goldman-Rakic, 1988).
Elektrische stimulatie van de hippocampus bij proefpersonen laat een dissociatieve symptomatologie zien met een subjectieve sensatie van angst, complexe visuele hallucinaties, flash backs, en emotionele onrust. Een bepaalde omgeving waarin de oorspronkelijke angstconditionering plaatsvindt kan een belangrijke rol spelen bij reacties op een volgende CS-aanbieding. De respons is des te sterker als de CS in de oorspronkelijke context wordt aangeboden. Contextuele conditionering wordt niet door de amygdala gemoduleerd maar zij wordt juist sterk gereguleerd door de hippocampus. Een laesie in de hippocampus voor een leertaak verhindert het optreden van contextuele conditionering, zonder dat dit overigens effect op de CS zelf heeft. De retrograde amnesie die optreedt voor contextuele angst is temporeel bepaald: een laesie tot twee weken na de leertaak levert nog retrograde amnesie op voor contextuele angst.
Onderzoeksbevindingen tonen een essentiële rol aan voor de hippocampus in de formatie van en consolidatie van declaratieve expliciete herinneringen in het geheugen, echter niet over langere duur. De rol bij de evaluatie van contextuele triggers is consistent met theorieën van de laatste jaren over spatiële, configuratieve en relationele verwerking (tijd-ruimteverankering).
Neurobiologische reacties op traumatische stress
De basis voor neurobiologische reacties op stress wordt vroeg in de ontwikkeling gelegd. Experimenteel onderzoek wijst uit dat ratten die neonataal een endotoxine kregen toegediend waarop zij reageerden met een fysiologische stressrespons, op volwassen leeftijd een grotere ACTH en corticosteroïderespons op stress lieten zien dan ratten uit een controlegroep. Daarnaast werd bij de experimentele groep een afname in receptordichtheid voor glucocorticoïden in verschillende hersenregionen gevonden (Shanks et al., 1995; Rots et al., 1996). Deze gegevens laten zien dat blootstelling aan endotoxine in een kritische ontwikkelingsfase de HPA-as verandert, met een verminderde negatieve feedback van ACTH op glucocorticoïde-inhibitie, wat de organismen vatbaar maakt voor met stress samenhangende ziekte. De basis voor sensitisering van met stress samenhangende neurobiologische systemen wordt aldus in een zeer vroege ontwikkelingsfase gelegd. Iedere opeenvolgende stressor grijpt in op de ingeslepen sensitisatie en op de ontstane allostatic load.
De arousal in relatie tot anticipatie of in antwoord op bedreiging kan een situatie doen ontstaan, waarvoor een aanzienlijke prijs betaald wordt. Het kan een neurale en hormonale activiteit opleveren die het vermogen om opnieuw te reageren op acute stressvolle veranderingen beperkt. Bij patiënten met PTSS kan bijvoorbeeld een dergelijk allostatisch evenwicht gevonden worden in het systeem van de HPA-as, waar hyperalertheid en vaak voorkomende prikkelbaarheid en irritatie samen leiden tot chronisch verhoogde spiegels van catecholaminen (Yehuda et al., 1996).
Op biologisch niveau zijn bij patiënten met PTSS verschillende veranderingen waargenomen: onder andere toegenomen glucocorticoid-receptoren op lymfocyten, toegenomen corticotropin realeasing hormone (CRH), toegenomen functie van HPA-as, fysiologische veranderingen op omgevingsstimuli, veranderingen in rapid eye movement (REM) densiteit, verminderde 24-uurs uitscheiding van cortisol, veranderde alfa-2-adrenerge receptor-binding, veranderde ACTH-respons op CRH enzovoort. Neurale en neuro-endocriene systemen zijn betrokken bij voortdurende stress of bij de effecten van acute stress. Op verschillende locaties treden veranderingen op: in het immuunsysteem (virale infecties; verhoogde metastatische spreiding van kanker), cardiovasculaire systeem (plaque-formatie die leidt tot atherosclerosis, plaque-rupture en tromboseneiging die leiden tot myocard-infarcering) vetweefsel en spieren (depositie van vet, verhoogde insuline secretie, samen leiden tot verhoogde atherosclerosis), en ook in het brein (McEwen, 1995).
Hoe zijn deze bevindingen terug te voeren tot de individuele patiënt? De klassieke patiënt met PTSS vertoont woede en agressie, angst, depressie, overeten, drankgebruik, roken, slechte spijsvertering en verhoogde hartslag; allemaal kenmerken van een slechte gezondheid. Sommigen lijden onder cardiovasculaire reactiviteit, toegenomen autonome arousal, verstoorde slaap, adrenerge disregulatie, verhoogde schildklierfunctie of een veranderde HPA-as activiteit. Uit epidemiologisch onderzoek blijkt dat blootstelling aan traumatische stress vaker leidt tot lichamelijke klachten, ziekte, en frequenter bezoek aan de dokter (Friedman & Schnurr, 1995).
Psychobiologische mechanismen die betrokken zijn bij de uitwerking van traumatische stress op geheugenopslag en ‘verwerking’
De evolutie heeft ons als zoogdieren voorzien van een geheugensysteem dat bijdraagt aan overleving, onder andere door de herkenning van gevaarlijke of bedreigende stimuli te vertalen via een hormonale respons. Dit neurale systeem is vroeg in de fylogenese ontstaan en is in essentie ongewijzigd gebleven tijdens de evolutie (McGaugh et al., 1992; McGaugh en Cahill, 1997). Bij de mens gaat het om overdracht van informatie via parallelle en complementaire kanalen en baansystemen naar de amygdala. De amygdala heeft als ‘taak’ deze informatie te integreren en zonodig een aantal typische overlevingsreacties tot stand te brengen. Het systeem kan slecht aangepast geraken en gaan dysfunctioneren, bijvoorbeeld als de balans tussen de deelnemende componenten verloren gaat. Dit kan optreden bij situaties van extreme of repititieve stress. De psychobiologische of neurale mechanismen van stress-sensitisatie, angstconditionering en extinctie beschrijven de aard van de relatie tussen impliciete geheugenfuncties en traumatische stress (Charney et al., 1993).
Stress-sensitisatie
Herhaalde exposure aan een stressor resulteert in verhoogde responsiviteit op de volgende stressoren. Acute stress veroorzaakt een toegenomen release van noradrenaline in de hippocampus en andere hersenregionen. In dierstudies werd een verhoogde noradrenaline-release gezien, bij blootstelling aan herhaalde stress. Patiënten met PTSS hebben juist moeite met stress, zelfs eenvoudige stress, een conflict met een wederhelft of een aanrijding in het verkeer, kunnen situaties opleveren waarin zij totaal decompenseren. Stress-sensitisatie, gepaard gaande met toegenomen noradrenaline-release kan een mogelijke verklaring zijn. Traumatisering in kinder- of adolescentietijd, gemeten met een vragenlijst naar vroege traumatisering, geeft een verhoogde kans op PTSS, zoals gevonden in een retrospectieve studie bij oorlogsveteranen (Bremner & Brett, 1997). Sensitisatie door vroege stress kan de kwetsbaarheid doen toenemen voor de ontwikkeling van psychopathologie na herhaling of opeenvolging van stress. Een periode van blootstelling aan stress verhoogt het risico voor stress-symptomatologie bij herhaalde blootstelling.
Na extreme stress is er meestal een massale secretie van neurotransmitters, gevolgd door uitputting en tekort. Na herhaalde trauma-tisering worden deze hypersecretie en daarop volgende tekorten een geconditioneerde respons, waardoor ook receptorveranderingen plaatsvinden, die leiden tot overreacties in daarop volgende stress-situaties.
Angstconditionering
Een neutrale stimulus wordt gepaard met een aversieve stimulus als een elektrische schok. Door herhaaldelijke gelijktijdige aanbieding van bijvoorbeeld licht en de schok ontstaat een leerproces waarin het licht alleen leidt tot een angstrespons. De respons kan jaren blijven bestaan. Zoals besproken lijkt de amygdala een rol te spelen bij het emotionele geheugen, zoals gemanifesteerd wordt bij geconditioneerde responsen. Elektrische stimulatie van de amygdala geeft bij gezonde personen gevoelens van angst en activatie van een stress-respons, wat te zien is aan een toename van perifere catecholaminen. Een verhoogde schrikreactie is een van de symptomen bij PTSS. Een psychofysiologisch paradigma levert veel mogelijkheden om de emotionele respons in het laboratorium te testen. Er is een verhoogde hartslag en bloeddruk bij PTSS-patiënten op prikkels die herinneren aan het oorspronkelijk trauma (films, geluid, verhalen), op basis waarvan een geconditioneerde respons in gang gezet kan worden.
Verminderde extinctie van angstverhogende stimuli
Voor een goed begrip: extinctie moet niet gezien worden als een passief vergeten, maar als een actief proces, dat waarschijnlijk te maken heeft met opnieuw leren. Misschien is de metafoor uitdoving niet goed gekozen om dit misverstand uit de wereld te helpen. De prefrontale cortex speelt een belangrijke rol bij het proces van extinctie. Deze cortex is essentieel bij het opnemen van bekrachtigende externe (mogelijk ook interne) stimuli. Verondersteld wordt dat het effect van de prefrontale cortex te vinden is in projecties naar de amygdala, waar een blokkade van bepaalde receptoren in de amygdala (N-methyl-D-aspartaat, NMDA) interfereert met de uitdoving van geconditioneerde angst. De gedachte dat het om opnieuw leren zou gaan komt voort uit de vaststelling dat er een betrokkenheid is van (NMDA) receptor-afhankelijke synaptische plasticiteit.
Verminderde extinctie is ook een karakteristiek verschijnsel bij PTSS. Extinctie refereert aan een inhibitie van een geconditioneerde respons op prikkels die geassocieerd worden met een angstwekkende stimulus en die geleidelijk toeneemt bij het verdwijnen of vermindering van de oorspronkelijke angstaanjagende stimulus. Patiënten die misbruikt/mishandeld werden in hun jeugd, en als onderdeel daarvan bijvoorbeeld werden opgesloten in een kast, kunnen later iedere keer als zij in een donkere of kleine ruimte terechtkomen een angst ervaren, hoewel deze ‘cues’ niet werkelijk representatief zijn voor de bedreiging van hun persoon die plaatsgevonden heeft. Hierbij treedt neocorticale en orbitofrontale inhibitie van de amygdala op. Daarnaast is een verminderd basaal glucosemetabolisme gevonden in de temporale cortex bij patiënten met PTSS vergeleken met controlepersonen gemeten met PET. Gespeculeerd wordt dat de temporale neocortex (auditore neocortex) onvoldoende extinctie geeft.
Stress veroorzaakt morfologische veranderingen in het brein
Een van de effecten van stress is dat veranderingen in de cytoarchitectuur van de hippocampus optreden, die onder andere leiden tot veranderingen in de expliciete geheugenfuncties. In studies met apen die blootgesteld werden aan extreme stress (zeer slechte behuizing) werden bij autopsie beschadigingen gevonden van verschillende regionen van de hippocampus (Uno et al., 1989). De apen hadden typische, met te grote belasting samenhangende afwijkingen: vergrote bijnieren, maagzweren, en atherosclerotische laesies. Hoge spiegels van glucocorticoïden die vrijkwamen tijdens stress werden in verband gebracht met deze hippocampale beschadiging, waaronder dendritische atrofie en neuronaal celverlies, welk verschijnsel een verklaring kan zijn voor de geheugendefecten zoals ook bij patiënten met PTSS gevonden worden. Glucocorticoïden oefenen het toxisch effect uit door een verhoogde kwetsbaarheid van hippocampale neuronen voor endogeen vrijgekomen aminozuren. Recente studies lieten een volumeverschil zien van de hippocampus bij PTSS-patiënten, zowel een groep Vietnam-veteranen als een groep seksueel-getraumatiseerde mensen (Bremner et al., 1995c; Bremner et al., 1997a). In de laatstgenoemde studie betrof het een ‘gematchte’ controlestudie van 17 volwassen seksueel getraumatiseerde personen, en was het gemiddelde volumetrisch verschil 12%. Er was geen significant volumetrisch verschil in andere hersengebieden. Deze patiënten hadden in vergelijking met de controlegroep een lagere score op de Wechsler Memory Scale. Er waren verschillen in het korte-termijngeheugen tussen de patiënten en de ‘normale’ controlegroep. Deze bevindingen zijn gerepliceerd (Stein et al., 1997; Gurvits et al., 1996). Een groep vond dat de ernst van de PTSS proportioneel was aan de mate van hippocampaal volumeverlies. Een hippocampaal volumeverschil in vergelijking met ‘normale’ personen is ook gevonden bij andere diagnosen, zoals depressie en schizofrenie, waardoor de specificiteit van deze bevindingen minder sterk wordt.
Minder is bekend over veranderingen in de amygdala. Er zijn geen volumetrische verschillen in de amygdala bij PTSS-studies gevonden. Het is niet duidelijk of glucocorticoïden de expressie van peptiden in de amygdala reguleren. PET-studies laten zien dat de amygdala selectief betrokken blijkt bij versterkte geheugensporen in een emotionele arousal. De toegenomen geheugenopslag is gecorreleerd met een toegenomen glucosemetabolisme in de amygdala (Cahill, 1997). Deze bevinding werd gerepliceerd door Rauch et al. (1995). De vraag is of de amygdala bij dissociatieve processen minder actief is, en of er een ‘numbness’ is als gevolg van een eventuele adrenerge blokkade. Dit zou de hypothese ondersteunen dat emotionele verwerking waarbij de amygdala niet betrokken is inadequaat verloopt (McGaugh et al., 1996).
Geheugenprocessen in beeld
Lange tijd heeft in de cognitieve psychologie het informatieverwerkingsparadigma een cruciale rol gespeeld, zonder dat elementen uit de neurobiologie geïntegreerd werden. Neuro-imaging levert nieuwe inzichten op. Gelokaliseerde neurale activiteit kan gemeten worden via metabole activiteit in hersenregionen. Beeldvormend onderzoek bij patiënten met PTSS dragen bij tot hypothesevorming over de pathofysiologie van traumatische stress. Onderzoek wordt bijvoorbeeld gedaan met stoffen met een korte halfwaardetijd, zoals radioactief water, waardoor informatie kan verkregen worden over veranderingen op het moment van bijvoorbeeld hyperarousal of intrusie. Imaging met stoffen met een korte halfwaardetijd heeft hierbij als voordeel dat specifieke taken, met een snelle verandering in het hersenmetabolisme in het onderzoek kunnen worden opgenomen (Rauch & Renshaw, 1996). Het blijkt dat het dorsolaterale cortex als deel van de prefrontale cortex een rol speelt bij het expliciete geheugen. De prefrontale cortex speelt een rol bij zowel het expliciete geheugen als bij aandachtsfuncties. De prefrontale cortex wordt beschreven als het interface tussen exteroceptieve en interoceptieve gewaarwordingen, waar integratie van sensorische gewaarwordingen plaatsvindt. De pariëtale cortex speelt een rol bij het spatiële geheugen en de aandacht. Bij psychofysiologische arousal bij PTSS-patiënten blijkt dat er sprake is van (rechtszijdige) activatie van amygdala, hippocampus, en visuele associatieve cortex. Er is afname van activiteit in de (linker) gyrus prefrontalis inferior, en Broca. Dit suggereert een hyperfunctie voor die hersengebieden betrokken bij emotie, geheugen en visuele processen, alle cognitieve aspecten die geactiveerd zijn bij PTSS. De inhibitie van Broca kan het omzetten van het trauma in narratieve elementen negatief beïnvloeden (Rauch et al., 1996). Uit het yohimbine-onderzoek waarbij het noradrenerge systeem gestimuleerd werd en PTSS-symptomen opzettelijk veroorzaakt werden, bleek een relatieve toename in het hersenmetabolisme in verschillende corticale gebieden (Bremner et al., 1997b). Functionele beeldvormende technieken zijn erg kostbaar; er is veel meer kennis nodig voor de bevindingen therapeutisch toepasbaar zijn. De aandacht gaat met name uit naar symptoomprovocatiemodellen, en het koppelen van PET aan structurele Magnetic Resonance Imaging (MRI) om de lokalisatie van metabole veranderingen, en dus hersenactiviteit, te verbeteren (Vermetten & Bremner, in druk).
Neurobiologische mechanismen bij dissociatieve amnesie
Dissociatie op het moment van trauma wordt beschouwd als een beschermende factor tegen het bewust meemaken van afkeer, pijn en hulpeloosheid, maar dit voordeel op korte termijn levert mogelijk moeilijkheden op langere termijn op (Marmar et al., 1996). Deze lange-termijneffecten kunnen gevonden worden in het geheugen, in de geheugenopslag, en in de integratie van de geheugeninhoud (Thygesen, Hermann & Willanger, 1970; Bremner et al., 1993; Yehuda et al., 1995c). Lange-termijnveranderingen in neurobiologische en neuro-endocriene systemen kunnen met zich meebrengen dat bij herhaalde blootstelling aan trauma dissociatieve reacties optreden, dat deze reacties vaker optreden en dat er een verhoogd risico ontstaat met stress samenhangende psychopathologie (Bremner et al, 1995b). Dissociatieve amnesie kan een symptoom zijn van een pathologische respons op trauma (Spiegel, 1988), zij kan ook een voorloper zijn van psychopathologie op langere termijn (Bremner & Brett, 1997).
Al eerder werd gesteld dat de neurale basis voor dissociatieve toestanden gevonden kan worden in de neurobiologie van het geheugen, gezien de intieme relatie tussen dissociatie en veranderingen in geheugenprocessen. Consistent hiermee is de bevinding dat stimulering van de hippocampus en naburige structuren symptomen doet veroorzaken die sterke gelijkenis vertonen met dissociatie. In een overzicht van Krystal wordt beschreven hoe toediening van ketamine-hydrochloride, een non-competitieve antagonist van N-methyl-D-aspartaat (NMDA)-receptor, leidt tot een toename in dissociatieve symptomatologie, zoals gemeten met de Clinician Administered Dissociative Symptoms Scale (CADSS) (Bremner et al., 1998b), en tot een verslechtering op resultaten op een woordherinneringstest bij gezonde proefpersonen. De NMDA-receptor, die in hoge concentratie voorkomt in de hippocampus, is betrokken bij geheugenopslag op moleculair niveau door een proces van lange-termijn-potentiëring (long term potentiation, LTP). Proefpersonen die ketamine namen beleefden een scala aan dissociatieve symptomen, waaronder ‘out-of-body’-ervaringen, het gevoel alsof hun armen stokjes waren, gaten in de tijdsbeleving, het gevoel alsof de tijd stil stond, verstoringen in het zelf-gevoel en derealisatie. Als deze experimenten vertaald worden naar het functioneren van de hippocampus levert dit een beeld op waarin dysfunctie van de hippocampus leidt tot een verlies van normale integratie van geheugen en bewustzijn. Deze dysfunctie omvat verstoringen in alle processen die betrokken zijn bij het geheugen: codering, consolidering, opslag of onderdelen van deze drie.
Dissociatie op het moment van psychologisch trauma kan een teken zijn van pathologische processen in onder andere de amygdala en hippocampus. Traumatische gebeurtenissen die aan het geheugen aangeboden worden op het moment dat een individu zich in een dergelijke toestand bevindt worden ook beter teruggehaald als de persoon zich in verwante toestand bevindt. Deze theorie ‘verklaart’ het fenomeen van de traumatische herbeleving.
Dissociatieve stoornissen passen in een neurobiologisch model over de invloed van stress op het geheugen. Geheugen kan ook in een ruime zin gedefinieerd worden, waarbij het autobiografisch geheugen bijdraagt tot de vorming van identiteit. Identiteit kan immers beschouwd worden als een serie van geheugeninhouden die iemands levensverhaal vormen. Identiteit is ‘the center of narrative gravity’ (Dennet, 1991). Traumatische stress wordt verondersteld de rode draad in het verhaal te verstoren. Dissociatieve symptomen van amnesie en identiteitsverstoring kunnen ook gerelateerd zijn aan de veranderde geheugenfunctie bij getraumatiseerde mensen.
Het belang van toestandsafhankelijkheid
Het terughalen van traumatische herinneringen vindt vaker plaats in een toestand die overeenkomt met de toestand waarin het trauma plaatsvond (Bower, 1981; Spiegel, 1990; Van der Kolk et al., 1996).
De literatuur is niet onverdeeld duidelijk in het geven van een verklaring voor het optreden van amnestische fenomenen op basis van dissociatie. Argumentatie voor dissociatieve momenten wordt gehaald uit een neurobiologische ‘state-dependency’-theorie. De theorie beschrijft hoe affectieve input bijdraagt tot overeenkomsten tussen opslag en teruggehaalde geheugeninhouden. De begrippen ‘state dependency’ of ‘mood-state-dependency’ geven aan hoe psychologische en biochemische geheugenfragmenten verankerd kunnen zijn, en op welke wijzen deze fragmenten specifieke bijdragen aan een geheugensysteem opleveren (Ellis, 1985; Kenealy, 1997). ‘Triggering’ van een enkel element kan een cascade van geheugeninhouden in het expliciete geheugen alsook geconditioneerde reacties oproepen.
Bepaalde herinneringen zijn met name of juist alleen bereikbaar in bepaalde (stemmings)toestanden, zoals agitatie of depressie. Afzonderlijke systemen voor de opslag van impliciete emotionele herinneringen en expliciete herinneringen voor emoties beïnvloeden de inhoud van de herinnering, zulks afhankelijk van emotionele toestanden. Om een herinnering bewust te laten worden moet het associatieve neurale netwerk een bepaald activatieniveau bereiken, afhankelijk van het aantal geheugencomponenten en het gewicht van elke van deze componenten. Hoe meer ‘cues’ van het leren die aanwezig zijn tijdens het herinneren, hoe zwaarder het gewicht van deze componenten, en des te waarschijnlijker zal deze herinnering optreden. Herinnering wordt bevorderd als de stemmingstoestand overeenkomt met die waarin de inhoud gecodeerd werd, maar zij kan ook worden gefaciliteerd door een specifieke stimulus (‘cue-dependency’).
Dat het geheugen vooral functioneert via dergelijke meer impliciete inhouden wordt te meer aannemelijk daar het expliciete geheugen nog niet volledig ontwikkeld is in kindertijd. Herinneringen aan extreme traumata zijn namelijk beter opgeslagen in het impliciete geheugen. Deze emotionele geheugeninhouden zijn met name toegankelijk voor het bewustzijn tijdens specifieke affectief beladen situaties, analoog aan het ‘state-dependent memory system’ (LeDoux, 1996).
Een mogelijke verklaring voor de affectieve ‘numbness’, zoals vaak bij dissociatieve stoornissen gezien wordt, kan gelegen zijn in een inhibitie van de noradrenerge arousal waardoor de activiteit in de amygdala, locus coeruleus en andere limbische structuren die betrokken zijn bij affectieve arousal verminderd wordt (LeDoux, 1996; Spiegel, 1997).
Het is ook mogelijk dat in een verhoogde arousal een ‘cue’ of een associatief neuraal netwerk initieert waardoor een cascade van geconditioneerde angstresponsen met dissociatieve symptomen optreedt. Een voorbeeld is de Vietnam-veteraan die jaren na terugkeer uit de oorlog winkelt met zijn vrouw. Hij heeft nog lange tijd na zijn terugkeer typische PTSS klachten. Deze zijn inmiddels in remissie. Hij is verder gezond, en leidt een betrekkelijk normaal sociaal leven. Het is druk in het winkelcentrum, waardoor hij enigszins gespannen raakt. Als hij een roltrap opgaat komt een Hindoestaanse vrouw de roltrap naast hem naar beneden. Het zien van deze vrouw zet bij de man een paniek in de gang, hij krijgt last van derealisatie en waant zich in Vietnam. Pas na enige tijd wordt duidelijk wat de ‘cue’ was: de vrouw had een stip op haar voorhoofd, en dit deed hem zich de laatste aanblik van een lotgenoot in Vietnam herinneren: een schotwond precies in het voorhoofd. Deze visuele stimulus tijdens hyperarousal gaf een cascade aan gefragmenteerde geheugeninhouden en reacties die leidden tot re-actualisering van zijn Vietnam-verleden, met een terugkeer van zijn PTSS-symptomen (Bremner, persoonlijke mededeling).
Therapeutische mogelijkheden voor symptomen van dissociatie die samenhangen met trauma
De dissociatieve stoornis is de laatste jaren goed beschreven, het is mogelijk de stoornis vast te stellen, te karakteriseren en de ernst in maat en getal uit te drukken (Spiegel & Cardena, 1991; Bremner, 1998). Het belang van psychotherapie bij de behandeling van dissociatieve stoornissen en de posttraumatische stress-stoornis is uiteraard groot. Diverse strategieën zijn beschreven om de invloed van vroegere traumata te verminderen, door zowel het trauma als de betekenis ervan in een geïntegreerde context te plaatsen (zie onder andere Putnam, 1989b; Van der Hart, 1995). Gegeven de gefragmenteerde sensorische natuur van gedissocieerde traumatische herinneringen waar geen ‘verhaal’ van gemaakt is, en waar geen verankering in ruimte en tijd voor gevonden is, is het van belang een procedure te vinden waarin de herinneringen wel verankerd kunnen worden. Dit zou een essentieel onderdeel zijn van psychotherapie, waarbij niet zozeer de catharsis wordt nagestreefd, maar meer het transformeren van sensorische ervaringen in woorden, om daarmee affecten, beelden en andere fragmenten van een nieuwe context te kunnen voorzien (Van der Kolk, 1994; Van der Kolk et al., 1996).
Ondanks wat bekend is over neurobiologische achtergronden van de dissociatieve stoornis en PTSS is er een opvallend gebrek aan meer dan puur symptoomgerichte farmacotherapeutische mogelijkheden. Ondanks een snel groeiende neurobiologische modelvorming over dissociatie zijn er nog weinig placebo-gecontroleerde farmacotherapeutische trials voor dissociatieve stoornissen verricht, en is er bijvoorbeeld geen specifieke antidissociatieve farmacotherapie ontwikkeld. Dit in contrast met ontwikkelingen op gebied van anxiolytica, antipsychotica, stemmingsstabilisatoren en antidepressiva. Er is echter enig onderzoek dat mogelijke aangrijpingsgebieden biedt voor farmacotherapie:
- Provocatiestudies met yohimbine en ketamine leveren informatie over de betrokken neurobiologische systemen en transmitters bij dissociatie. Stoffen worden ontwikkeld om zo de basale desintegratie in de traumatische ervaring te behandelen. Hierbij lijkt vooral de NMDA-receptor van betekenis (Krystal et al., 1998).
- Als een irreversibele toename in excitatie optreedt bij PTSS ligt het meer voor de hand een strategie te zoeken om PTSS te voorkomen dan PTSS te behandelen. Suppressie van de b-adrenerge activiteit voorafgaand of juist na een traumatische gebeurtenis wordt als mogelijkheid genoemd (Cahill, 1997). Dit zou concreet betekenen dat het therapeutisch kan zijn om iemand direct na een doorgemaakt trauma een b-blokker te geven.
- Een andere farmacotherapeutische ingang zou gericht kunnen zijn op het tegengaan van veranderingen in de cytoarchitectuur en het neuraal verval van de hippocampus. Gedacht wordt aan cyclisch adenosine 3,5-fosfaat (cAMP), als ‘second messenger’ bij de intracellulaire-overdracht van informatie en ‘brain derived neurotrofic factor’ (BDNF), een stof betrokken bij de nieuwvorming van dendritische connecties. Dit zijn stoffen waarnaar ook in het kader van depressieve stoornissen veel onderzoek plaatsvindt (Dunman et al., 1997).
Afsluiting
Traumatische stress kan in een grote variëteit aan symptomen resulteren, waaronder lange-termijneffecten op hersensystemen die bij het geheugen betrokken zijn. In dit artikel is getracht een overzicht te geven van de gevolgen van stress voor het individu, waarbij benadrukt is dat psychisch trauma samenhangt met een verhoogde symptomatologie van PTSS en dissociatieve stoornissen. Veranderingen in hersenregionen die betrokken zijn bij het geheugen kunnen een verband vormen tussen dissociatie en andere symptomen van traumatische stress. Stress geeft aanleiding tot veranderingen in het brein die op langere termijn manifest worden. Er zijn aanwijzingen dat dysfuncties in deze hersenregionen de symptomen van dissociatie en PTSS kunnen mediëren. Het lijkt in de huidige neurobiologische modelvorming moeilijk om fenomenen van fictieve of geïmplanteerde herinneringen te verklaren. Het feit dat er bij traumatische herinneringen plastische veranderingen met neuro-endocriene parameters opgetreden zijn, maakt eenzelfde impact bij niet doormaakte traumata onwaarschijnlijk. Een goed begrip van de effecten van stress op het geheugen en de relatie tussen veranderingen in het geheugen en dissociatieve stoornissen kan een aanzet bieden voor de ontwikkeling van nieuwe therapeutische mogelijkheden bij de behandeling van ernstig getraumatiseerde patiënten op zowel psychotherapeutisch als biologisch gebied.
Dankbetuiging
Dank aan Onno van der Hart en Thea Overbeek voor hun bijdrage in de totstandkoming van dit artikel.
Summary
Currently there is a strong interest in the relationship between stress and memory. In an actual discussion on the relation between trauma, dissociation and trauma, knowledge of current neurobiological research is essential for a thorough interpretation of the phenomena of dissociation and posttraumatic stress. Traumatic stress results in changes in brain regions involved in memory functions that may effect the response; changes in these regions can play a role in the mediation of phenomena of dissociation and posttraumatic stress. Neuroendocrine and neuroanatomical limbic structures are described as central elements involved in the integration of declarative and implicit neurobiological functions. Neural mechanisms as fear conditioning, stress sensitization, and extinction are involved in the effects of stress on memory. The concept of neural associative networks is supportive of a state-dependency-explanation for dissociative phenomena in the background of encoding, storage and retrieval of memory fragments. The fact that in traumatic memories plastic changes with neuroendocrine parameters are involved, makes the same impact in false or implanted memories doubtful. Possible anti-dissociative agents in treatment of dissociative disorders ares described. Current research shows that stress induced alternations in brain regions are involved in memory support dissociative amnestic phenomena and symptoms of post traumatic stress.
Referenties
American Psychiatric Association. (1994). Diagnostic and Statistical Manual of Mental Disorders (Fourth Edition). Washington, DC: American Psychiatric Association.
Appelbaum, P.S., Uyehara, L.A., en Elin, M.R. (red.) (1997). Trauma and Memory; Clinical and Legal Controversies, New York: Oxford University Press.
Barnier, A.J. & McConkey, K.M. (1992). Reports of real and false memories: the relevance of hypnotizability and context of memory test. Journal of Abnormal Psychology, 101, 3, 521-527.
Boon, S. & Draijer, N. (1993). Multiple personality disorder in the Netherlands: A study on reliability and validity of the diagnosis. Amsterdam: Swets en Zeitlinger.
Bower, G.H. (1981). Mood and memory. American Psychologist, 36, 129-148.
Bremner, J.D., Southwick, S., Brett, E., Fontana, A., Rosenheck, R. & Charney, D.S. (1992). Dissociation and posttraumatic stress disorder in Vietnam combat veterans. American Journal Psychiatry, 149, 328-333.
Bremner, J.D., Davis, M., Southwick, S.M., Krystal, J.H. & Charney, D.S. (1993). Neurobiology of posttraumatic stress disorder. In : J.M. Oldham, M.B. Riba, and A. Tasman (red.), Review of Psychiatry, volume 12 (p. 183-205). Washington DC: American Psychiatric Press.
Bremner, J.D., Randall, P., Scott, T., Capelli, S., Delaney, R., McCarthy, G. & Charney, D.S. (1995a). Deficits in short-term memory in adult survivors of childhood abuse. Psychiatry Research, 59, 97-107.
Bremner, J.D., Krystal, J.H., Southwick, S.M., & Charney, D.S. (1995b). Functional Neuro-anatomical Correlates of the Effects of Stress on Memory. Journal Traumatic Stress, 8, 527-553.
Bremner, J.D., Randall, P., Scott, T.M., Bronen, R.A., Seibyl, J.P., Southwick, S.M., Delaney, R.C., McCarthy, G., Charney, D.S. & Innis, R.B. (1995c). MRI-based measurements of hippocampal volume in combat related posttraumatic stress disorder. American Journal Psychiatry, 152, 973-981.
Bremner, J.D., Krystal, J.H., Charney, D.S., & Southwick, S.M. (1996). Neural mechanisms in dissociative amnesia for childhood abuse: relevance to the current controversy surrounding false memory syndrome. American Journal Psychiatry, 8, 71-82.
Bremner, J.D. & Brett, E. (1997). Trauma-related dissociative states and long-term psychopathology in posttraumatic stress disorder. Journal Traumatic Stress, 10 (1), 37-51.
Bremner, J.D., Randall, P., Vermetten, E., Staib, L., Bronen, R.A., Mazure, C., Capelli, S., McCarthy, G., Innis, R. & Charney, D.S. (1997a). Magnetic resonance imaging-based measurement of hippocampal volume in posttraumatic stress disorder related to childhood physical and sexual abuse; a preliminary report. Biological Psychiatry, 41, 23-32.
Bremner, J.D., Innis, R.B., Ng, C.K., Staib, L.H., Salomon, R.M., Bronen, R.A., Duncan, J., Southwick, S.M., Krystal, J.H., Rich, D., Zubal, G., Dey, H., Soufer, R. & Charney, D.S. (1997b). Positron Emission Tomography Measurement of Cerebral Metabolic Correlates of Yohimbine Administration in Combat-Related Posttraumatic Stress Disorder. Archives General Psychiatry, 54, 246-254.
Bremner, J.D., Vermetten, E., Southwick, S.M. Krystal, J.H. & Charney, D.S. (1998a). Trauma, Memory and Dissociation, : An Integrative Formulation In: J.D. Bremner & C. Marmar (red.) Trauma, Memory and Dissociation. Washington, DC: American Psychiatric Association Press.
Bremner, J.D., Krystal, J.H., Putnam, F.W., Southwick, S.M., Marmar, C., Charney, D.S., Mazure, C.M. (1998b). Measurement of dissociative states with the clinician-administered dissociative states scale (CADDS). Journal Traumatic Stress, 11 (1), 125-137.
Brewin, C.R. (1996). Scientific Status of Recovered Memories. British Journal of Psychiatry, 131-134.
Briere, J. & Conte, J. (1993). Self-reported amnesia for abuse in adults molested as children. Journal Traumatic Stress, 6 (1), 21-31.
Brown, D., Scheflin, A.W. & Hammond, C. (1998). Memory, Trauma Treatment and the Law. New York: W.W. Norton and Company.
Brown, K. & Kulik J. (1977). Cognition, 5, 73-99.
Brown, D. (1995). Pseudomemories: the standard of science and the standard of care in trauma treatment. American Journal Clinical Hypnosis, 37, 3, 1-24.
Cahill, L., Prins, B., Weber, M. & McGaugh, J.L. (1994). B-adrenergic activation and memory for emotional events. Nature, 371 (20), 702-704.
Cahill, L. & McGaugh, J.L. (1995) A novel demonstration of enhanced memory associated with emotional arousal. Consciousness and Cognition, 4, 41-42.
Cahill, L. (1997). Neurobiology of emotionally influenced memory. In: R.Yehuda & A.C. McFarlane (red.), Psychobiology of Post Traumatic Stress Disorder. Volume 821 (p. 238-246). New York: Annals of the New York Academy of Sciences.
Carlier, I.V., Lamberts, R.D., Fouwels, A.J. & Gersons, B.P. (1996). PTSD in relation to dissociation in traumatized police officers. American Journal Psychiatry, 153 (10), 1325-1328.
Charney, D.S., Deutch, A.Y., Krystal, J.H., Southwick, S.M. & Davis M. (1993). Psychobiologic mechanisms of posttraumatic stress disorder. Archives of General Psychiatry, 50, 294-305.
Christianson, S.-A. & Loftus, E. (1987). Memory for traumatic events. Applied Cognitive Psychology, 1, 225-239.
Christianson, S.-A., & Engelberg, E. (1997). Remembering and forgetting traumatic experiences: a matter of survival. In: M.A. Conway (red.), Recovered Memories and False Memories, (p. 230-251), Oxford: Oxford University Press.
Chrousos, G.P. & Gold, P.W. (1992). The concepts of stress and stress system disorders: Overview physical and behavioral homeostasis. Journal of the American Medical Association, 267, 1244-1252.
Chu, J.A. & Dill, D.L. (1990). Dissociative symptoms in relation to childhood physical and sexual abuse. American Journal Psychiatry, 147, 887-892.
Conway, M.A., Anderson, S.J., Larsen, S.F., Donnelly, C.M., McDaniel, M.A., McClelland, A.G.R., Rawles, R.E. & Logie, R.H. (1994). Memory and Cognition, 22, 326-343.
Coons, P.M. (1994). Confirmation of childhood abuse in child and adolescent cases of multiple personality disorder and dissociative disorder not otherwise specified. Journal Nervous and Mental Disorders, 8, 461-464.
Crombag, H.F.M. & Merckelbach, H.L.G.J. (1996). Hervonden herinneringen en andere misverstanden. Amsterdam, Contact.
Dennett, D. (1991). Consciousness Explained. Boston: Little Brown.
Draijer, N. (1997).Hervonden misdrijven eenzijdig bekeken. Maandblad Geestelijk Volksgezondheid, 12, 1249-1256.
Dunman, R.S., Heninger, G.R., & Nestler, E.J. (1997) A molecular and cellular theory of depression. Archives of General Psychiatry, 7
Edelman, G.M. (1989). Basic Books, New York.
Eich, J.E. (1977). State-dependent memory retrieval of information in human episodic memory. In I.M. Birnbaum & E.S. Parker (red.). (p. 141-157), Hillsdale, NJ: Erlbaum.
Eich, J.E. (1980). The cue-dependent nature of state-dependent retrieval. Memory and Cognition, 8, 2, 157-173.
Ellis, M.E. (1985). Amygdala norepinefrine involved in two separate long-term memory retrieval processes. Brain Research, 342 (1), 191-195.
Erdelyi, M.H., (1996). The Recovery of Unconscious Memories; Hypermnesia and Reminiscence. Chicago: University of Chicago Press.
Eriksson, N.G., & Lundin, T. (1996). Early traumatic stress reactions among Swedish survivors of the Estonia disaster. British Journal Psychiatry, 169, 713-716.
Friedman, M.J., Charney, D.S. & Deutch, A.Y. (1995). Neurobiological and Clinical Consequences of Stress: From Normal Adaptation to Post Traumatic Stress Disorder. Philadelphia: Lippincot.
Friedman, M.J. & Schnurr, P. (1995). The Relationship between trauma, post-traumatic stress disorder, and physical health. In: M.J. Friedman, Charney D.S. & Deutch, A. Neurobiological and Clinical Consequences of Stress. (p. 507-527). Philadelphia: Lippincott-Raven.
Godden, D.R. & Baddeley, A.D. (1975). Context-dependent memory in two natural environments: On land and under water. British Journal Psychology, 66, 325-331.
Gold, P.E. & McGaugh, J.L.(1977). In: D. Deutch, & J.A. Deutch (red.), Short term memory. (p. 355-378). New York: Academic Press Inc.
Goldman-Rakic, P.S. (1988). Circuitry of primate prefrontal cortex and regulation of behavior by representational memory. In F. Plum (red.), Handbook of physiology, section 1: higher functions of the brain. (p. 373-417). New York: Oxford University Press.
Gurvits, T.V., Shenton, M.E., Hokama, H., Ohta, H., Lasko, N.B., Gilbertson, M.W., Orr, S.P., Kikinis, R., Jolesz, F.A., McCarley, R.W. & Pitman, R.K. (1996). Magnetic resonance imaging study of hippocampal volume in chronic, combat-related posttraumatic stress disorder. Biological Psychiatry, 40, 1091-1099.
Hammond, C.D., Garver, R.B., Mutter, C.B., Crasilneck, H.B., Frischholz, E., Gravitz, M.A., Hibler, N.S., Olson, J., Scheflin, A., Spiegel, H. & Wester, W. (1995). Clinical Hypnosis and memory: guidelines for clinicians and for forensic hypnosis. American Society of Hypnosis Press.
Herman, J. (1992). Trauma and Recovery. New York: Basic Books.
Hart, O. van der (red.) (1995). Trauma, dissociatie en hypnose. Lisse: Swets en Zeitlinger.
Hopfield, J.J. (1982). Neural networks and physical systems with emergent collective computational abilities. Proceedings of the National Academy of Science USA, 79, 2554-2558.
Irwin, H.J. (1994). Proneness to dissociation and traumatic childhood events. Journal Nervous and Mental Disorders, 8, 456-460.
Jeffery, K.J. & Reid, I.C. (1997). Modifiable neuronal connections: an overview for psychiatrists. American Journal of Psychiatry, 154 (2), 156-164.
Kandel, E.R. & Schwartz, J.H. (1985). Principles of Neural Science. New York: Elsevier.
Kenealy, P.M. (1997). Mood-state-dependent retrieval: the affects of induced mood on memory reconsidered. Quarterly Journal of Experimental Psychology A, 50, 2, 290-317.
Kihlstrom, J.F. (1997). Hypnosis memory and amnesia. Philos Trans R Soc Lond B Biol Sci, 352, 1362, 1727-1732.
Kolk, B.A. van der, Hart, O. van der, & Marmar, Ch. (1996). Dissociation and Information Processing in Posttraumatic Stress Disorder. In: B.A. Van der Kolk, A. McFarlane, L. Weisaeth (red.). Traumatic Stress; the Effects of Overwhelming Experience on Mind, Body and Society. Pp. 303-331. New York: Guilford.
Kolk, B.A. van der, Burbridge, J.A. & Suzuki, J. (1997).The psychobiology of traumatic memory. Clinical implications of neuroimaging studies. In: Yehuda, R. & McFarlane, A.C. (red.), Psychobiology of Posttraumatic Stress Disorder, (p. 99-114). New York: Annals of the New York Academy of Sciences, vol 821.
Kolk, B.A. van der & Hart, O. van der (1989). Pierre Janet and the breakdown of adaptation in psychological trauma. Am J of Psych. 146, 12, 1530-1540.
Kolk, B.A. van der (1994). The body keeps the score: memory and the evolving psychobiology of posttraumatic stress. Harvard Rev Psychiatry, 253-265.
Krystal, J.H., Bennett, A., Bremner, J.D., Southwick, S.M. & Charney, D.S. (1995). Toward a cognitive neuroscience of dissociation and altered memory functions in post-traumatic stress disorder. In: M.J. Friedman, D.S. Charney & A.Y. Deutch (red.). Neurobiological and Clinical Consequences of Stress: From Normal Adaptation to Post Traumatic Stress Disorder, (p. 239-271). Philadelphia: Lippincot.
Krystal, J.H., Bremner, J.D., Soiuthwick, S.M., Charney, D.S., (1998). The emerging neurobiology of dissociation: implications for the treatment of posttraumatic stress disorder. In: J.D. Bremner & Marmar Ch., (red.), Trauma, memory and dissociation. (p. 321-363). Washington, DC: American Psychiatric Press.
Labelle, L., Laurence, J.R., Nadon, R. & Perry, C. (1990). Hypnotizability preference for an imaginative cognitive style and memory creation in hypnosis. Journal of Abnormal Psychology, 99 (3), 222-228.
Leavitt, F. (1997). False attribution of suggestibility to explain recovered memory of childhood sexual abuse following extended amnesia. Child Abuse & Neglect, 21 (3), 265-272.
LeDoux, J.E. (1992). Emotion as Memory: Anatomical systems underlying indelible neural traces. In. A. Christianson (red.) Handbook of Emotion and Memory. (p. 269-288). Hillsdale, N.J.: Lawrence Erlbaum.
LeDoux, J.E. (1996). The Emotional Brain: The Mysterious Underpinnings of Emotional Life. New York, Simon and Schuster.
Loewenstein, R.J. (1993). Dissociation, development and the psychobiology of trauma. Journal American Academia Psychoanalysis, 21, 581-603.
Loftus, E.F. (1994). The repressed memory controversy. American Psychologist, 49, 443-445.
Lynn, S.J. & Rhue, J.W. (1986). The fantasy prone personality: hypnosis, imagination and creativity. J Pers Soc Psychol, 51 (2), 404-408.
Lynn, S.J. & Rhue, J.W. (1988). Fantasy Proneness: Hypnosis, developmental antecedents, and psychopathology. American Psychologist, 43, 35-44.
Marmar, C.R., Weiss, D.S. & Schlenger, D.S. (1994). Peritraumatic dissociation and posttraumatic stress in male Vietnam Theater veterans. American Journal of Psychiatry, 151, 902-907.
Marmar, C.R., Weiss, D.S., Metzler, T.J. & Delucci, K. (1996). Characteristics of emergency personnel related to peritraumatic dissociation during critical incident exposure. American Journal of Psychiatry, 153, 7, suppl, 94-102.
Mason, J.W., Giller, E.L. & Kosten, T.R. (1986). Urinary-free cortisol levels in post-traumatic stress disorder patients. Jourrnal of Nervous and Mental Disorders, 174, 145-159.
Mazure, C.M. (red.) (1994). Stress and psychiatric disorders. Washington DC, American Psychiatric Press.
McCloskey, M.C., Wible, N. & Cohen, N.J. (1988). J Exp Psych Gen, 117, 336-338.
McEwen, B.S. & Schmeck, H.M. (1994). The Hostage Brain. Rockefeller University Press, New York.
McEwen, B.S. & Sapolski R.M. (1995). Stress and Cognitive Function. Curr Opinion Neurobiology, 5, 205-216.
McGaugh, J.L.,Introini-Collison, I.B., Cahill, L., Kim, M. & Liang, K.C. (1992). In: J.P. Aggleton (red.), The Amygdala: Neurobiological Aspects of Emotion, Memory, and Mental Dysfunction. (p. 431-451), New York: Wiley-Liss.
McGaugh, J.L., Cahill, L. & Roozendaal, B. (1996). Involvement of the Amygdala in Memory Storage: Interaction with other Brain Systems. Proceedings National Academy of Science
USA, 93, (24), 13508-13514.
McGaugh, J.L. & Cahill, L. (1997). Interaction of Neuromodulatory Systems in Modulating Memory Storage. Behavour Brain Research, 83, 1-2, 31-38.
Mishkin, M. (1978). Memory in monkeys severely impaired by combined but not by separate removal of amygdala and hippocampus. Nature, 273, 5660, 297-298.
Neisser, U. (1982). In: U. Neisser (red.) Memory Observed: Remembering in natural contexts. (p. 43-48). San Fransisco: Freeman.
Nemiah, J.C. (1989). Janet redivivus: the centenary of ‘l Automatisme Psychologique. American Journal of Psychiatry, 146, 12, 1527-1530.
Packard, M.G., Williams, C.L., Cahill, L. & McGaugh, J.L. (1995) In: N.E. Spear, L.P. Spear & M.L. Woodruff (red.), Neurobehavioral Plasticity: learning, development, and response to brain insults. (p. 149-184), New Jersey: Lawrence Erlbaum.
Parks, R.W., Long, D.L., Levine, D.S., Crockett, D.J., McGeer, E.G., McGeer, P.L., Dalton, I.E., Zec, R.F., Becker, R.E., Coburn, K.L., Siler, G., Nelson, M.E. & Bower, J.M. (1991). Parallel distributed processing and neural networks: origins, methodology and cognitive functions. International Journal of Neuroscience. 60, 195-214.
Pitman, R. & Orr, S. (1990). Twenty-four hour urinary cortisol and catecholamine excretion in combat-related PTSD. Biological Psychiatry, 27, 245-247.
Putnam, F.W. (1985). Dissociation as a response to extreme trauma. In: R.P. Kluft (red.). Childhood Antecedents of Multiple Personality Disorder, (p. 65-97). Washington, DC: American Psychiatric Press.
Putnam, F.W. (1989a). Pierre Janet and modern views of dissociation. Journal of Traumatic Stress, 2, 413-429.
Putnam, F.W. (1989b). Diagnosis and Treatment of Multilpe Personality Disorder. New York, Guilford.
Putnam, F.W. (1991). Dissociative Phenomena. In A. Tasman, & S.M. Goldfinger (red.). American Psychiatric Press Review of Psychiatry – Volume 10. (p. 145-160). Washington DC: American Psychiatric Press.
Rauch, S.L. & Renshaw, P.T. (1995). Clinical neuroimaging in psychiatry. Harvard Review of Psychiatry
2, 297-312.
Rauch, S.L., Van der Kolk, B.A., Fisler, R.E., Alpert, N.M., Orr, S.P., Savage, A.J., Fishman, M., Jenike, M.A. & Pitman, R.K. (1996). A symptom provocatio study of posttraumatic stress disorder using positron emission tomography and script driven imagery. Archives of General Psychiatry, 53, 380-387.
Ross, C.A., Joshi, S. & Currie, R.P. (1991). Dissociation in the general population: identification of three factors. Hospital Community Psychiatry. 42, 297-301.
Rots, N.Y., De Jong, N., Workel, J.O., Levine, S., Cools, A.R. & De Kloet, E.R. (1996). Neonatal maternally deprived rats have as adults elevated basal pituitary-adrenal activity and enhanced susceptibility to apomorphine. J Neuroendo. 8, 501-506.
Rumelhart, D.E. & McClelland, J.L. (1986). Parallel Distributed Processing: Explorations in the Microstructures of Cognition, Vol. 1.: Foundations. Cambridge, ma: mit Press.
Sapolski, R. (1996). Why stress is bad for your brain. Science,
273, 749-750.
Schacter, D.L., (1995) Implicit memory: a new frontier for cognitive neuroscience. In: Gazzinga, M.S., (red.), The cognitive neurosciences. Cambridge, ma, mit Press.
Schacter, D., (1996). Searching for memory; The Brain, the Mind and the Past. New York, Basic Books.
Scheflin, A.W. & Brown, D. (1996). Repressed memory or dissociative amnesia: what the science says. Journal of Psychiatry and Law, 2, 143-188.
Selye, H. (1936). A syndrome produced by diverse noxious agents. Nature, 138, 32-36.
Shanks, D.R. (1995). Is human learning rational? Journal of Experimental Psychology A, 48, 2, 257-279.
Southwick, S.M., Krystal, J.H., Morgan, A. & Charney, D.S. (1991). Yohimbine and m-chlorophenylpiperazine in PTSD, New Research Programs and Abstracts (NR 348). Washington DL, American Psychiatric Association.
Southwick, S.M., Bremner, J.D., Krystal, J.H. & Charney, D.S. (1994). Psychobiologic research in posttraumatic stress disorder. Psychiatr Clin North Am, 17, 251-264.
Southwick, S.M., Krystal, J.H., Morgan, A., & Charney, D.S. (1993). Abnormal adrenergic function in posttraumatic stress disorder. Archives of General Psychiatry, 50, 266-274.
Spiegel, D., Hunt, T. & Dondershine, H.E. (1988). Dissociation and Hypnotizability in post traumatic stress disorder. American Journal of Psychiatry, 145, 301-305.
Spiegel, D. (1990). Hypnosis, Dissociation and Trauma: Hidden and Overt Observers. In: J.L. Singer JL. (red.) Repression and Dissociation. (p. 121-142). Chicago: University of Chicago Press.
Spiegel, D. & Cardena, E. (1991). Disintegrated experience: The dissociative disorders revisited. Journal of Abnormal Psychology.
100, 366-378.
Spiegel, D. (1997) Trauma, Dissociation and Memory. In: Yehuda, R. & McFarlane, A.C. (red.), Psychobiology of Posttraumatic Stress Disorder, (p. 225-237). New York: Annals of the New York Academy of Sciences, vol 821.
Squire L.R. & Zola-Morgan, S. (1991). The medial temporal lobe memory system. Science, 153, 2380-2386.
Stein, M., Hanna, C., Koverola, C., Torchia, M & McClarty, B. (1997). Structural Brain Changes in PTSD. Does Trauma alter Neuroanatomy? In: Yehuda, R. & McFarlane, A.C. (red.),
Psychobiology of Posttraumatic Stress Disorder, (p. 76-83). New York: Annals of the New York Academy of Sciences, vol 821.
Stratton, G.M. (1919). Psychology Review, 26: 474-486.
Terr, L. (1991). Childhood traumas: An outline and overview. American Journal of Psychiatry, 27, 96-104.
Terr, L.C. (1996). Childrens memories in the wake of the Challenger. American Journal of Psychiatry, 153, 5, 618-25.
Thygesen, P., Hermann, K. & Willanger, R. (1970). Concentration camp survivors in Denmark: Prosecution, disease, disability, compensation. Danish Medical Bulletin, 17, 65-108.
Uno, H., Tarara, R., Else, J.G., Suleman, M.A. & Sapolsky, R.M. (1989). Hippocampal damage associated with prolonged and fatal stress in primates. Journal of Neuroscience, 9, 1705-1711.
Vanderwolf, C.H. & Cain, D.P. (1994). The behavioral neurobiology of learning and memory: a conceptual reorientation. Brain Research Review, 19, 3, 264-279.
Vermetten, E., Bremner, J.D. & Spiegel, D. (1998). Dissociation and Hypnotic Susceptibility: A Conceptual and Methodological Perspective on Two Distinct Concepts. In: J.D. Bremner & Ch. Marmar (red.). Trauma, Memory and Dissociation, pp 107-161. Washington, DC: American Psychiatric Press.
Vermetten, E. & Bremner, J.D. (in press). Dissociatieve Amnesia. In: G.E. Berrios & J. Hodges (red.) Memory Complaints and Disorders: The Neuropsychiatric Perspective. Cambridge: Cambridge University Press.
Vermetten E, Bremner JD. (in press) Anatomically defined regions of interest based on MRI-PET Cogegistration: Development, Rationale and Reliability of Method. Psychiatry Research.
Wakefield, H. & Underwager, R. (1992). Recovered memories of alleged sexual abuse: Lawsuits against parents. Behavioral Sciences and the Law, 10, 483-507.
Williams, L.M. (1994). What does it mean to forget child sexual abuse? A reply to Loftus, Garry and Feldman. Journal of Consulting and Clinical Psychology, 62, 1182-1186.
Williams, L.M. (1994b). Adult memories of childhood abuse. Journal of Consulting and Clinical Psychology, 62, 1167-1176.
Williams, L.M. (1995). Recovered memories of abuse in women with documented child sexual victimization histories. Journal of Traumatic Stress, 8, 649-676.
Yehuda, R., Kahana, R.B., & Binder-Brynes, K. (1995a). Low urinary cortisol excretion in holocaust survivors with PTSD. American Journal of Psychiatry, 152, 7-12.
Yehuda, R., Boisoneau, M.T., Lowy, M.T. & Giller, E.L. (1995b). Dose-response change changes in plasma cortisol and lymfocyte glucocorticoid receptors following dexamethasone administration in combat veterans with and without posttraumatic stress disorder. Archives of general Psychiatry, 52, 583-593.
Yehuda, R., Giller, E.L., Levengood, R.A., Southwick, S.M., & Siever, L.J. (1995c). Hypothalamic-pituitary-adrenal functioning in post-traumatic stress disorder: Expanding the concept of the stress response spectrum. In: M.J. Friedman, D.S. Charney & A.Y. Deutch (red.), Neurobiological and Clinical Consequences of Stress: From Normal Adaptation to PTSD. (p. 351-365). Philadelphia: Lippincot-Raven.
Yehuda, R., Teicher M.H., & Trestman R.L. (1997). Cortisol regulation in posttraumatic stress disorder and major depression: A chronobiological analysis. Biological Psychiatry, 40, 79-88.
Zola-Morgan, S., Squire, L.R. & Amaral, D.G. (1989). Lesions of perirhinal and parahippocampal cortex that spare the amygdala and hippocampal formation produce severe memory impairment. Journal of Neuroscience, 9, 4355-4370.